

The effect of the dominant polychaete Scolelepis squamata on ...
The effect of the dominant polychaete Scolelepis squamata on ...
The effect of the dominant polychaete Scolelepis squamata on ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
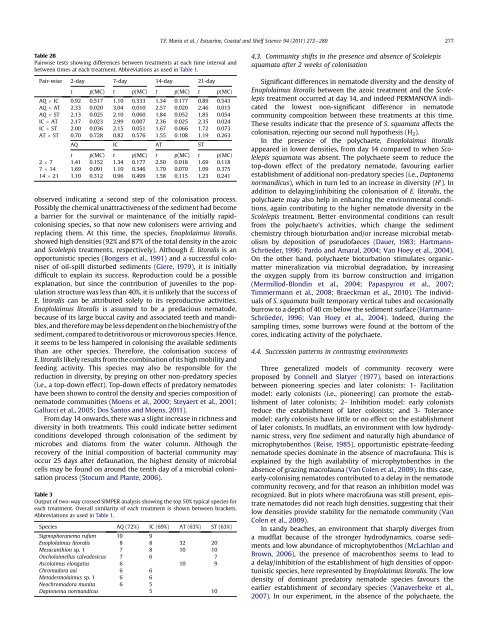
Table 2B<br />
Pairwise tests showing differences between treatments at each time interval and<br />
between times at each treatment. Abbreviati<strong>on</strong>s as used in Table 1.<br />
Pair-wise 2-day 7-day 14-day 21-day<br />
t p(MC) t p(MC) t p(MC) t p(MC)<br />
AQ IC 0.92 0.517 1.10 0.333 1.34 0.177 0.89 0.543<br />
AQ AT 2.33 0.020 3.04 0.010 2.57 0.020 2.46 0.015<br />
AQ ST 2.13 0.025 2.10 0.060 1.84 0.052 1.85 0.054<br />
IC AT 2.17 0.023 2.99 0.007 2.36 0.025 2.35 0.024<br />
IC ST 2.00 0.036 2.15 0.051 1.67 0.066 1.72 0.073<br />
AT ST 0.70 0.728 0.82 0.576 1.55 0.108 1.19 0.263<br />
AQ IC AT ST<br />
t p(MC) t p(MC) t p(MC) t p(MC)<br />
2 7 1.41 0.152 1.34 0.177 2.50 0.018 1.69 0.118<br />
7 14 1.69 0.091 1.10 0.346 1.79 0.070 1.09 0.375<br />
14 21 1.10 0.312 0.96 0.499 1.58 0.115 1.23 0.241<br />
observed indicating a sec<strong>on</strong>d step <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> col<strong>on</strong>isati<strong>on</strong> process.<br />
Possibly <str<strong>on</strong>g>the</str<strong>on</strong>g> chemical unattractiveness <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> sediment had become<br />
a barrier for <str<strong>on</strong>g>the</str<strong>on</strong>g> survival or maintenance <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> initially rapidcol<strong>on</strong>ising<br />
species, so that now new col<strong>on</strong>isers were arriving and<br />
replacing <str<strong>on</strong>g>the</str<strong>on</strong>g>m. At this time, <str<strong>on</strong>g>the</str<strong>on</strong>g> species, Enoplolaimus litoralis,<br />
showed high densities (92% and 87% <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> total density in <str<strong>on</strong>g>the</str<strong>on</strong>g> azoic<br />
and <str<strong>on</strong>g>Scolelepis</str<strong>on</strong>g> treatments, respectively). Although E. litoralis is an<br />
opportunistic species (B<strong>on</strong>gers et al., 1991) and a successful col<strong>on</strong>iser<br />
<str<strong>on</strong>g>of</str<strong>on</strong>g> oil-spill disturbed sediments (Giere, 1979), it is initially<br />
difficult to explain its success. Reproducti<strong>on</strong> could be a possible<br />
explanati<strong>on</strong>, but since <str<strong>on</strong>g>the</str<strong>on</strong>g> c<strong>on</strong>tributi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> juveniles to <str<strong>on</strong>g>the</str<strong>on</strong>g> populati<strong>on</strong><br />
structure was less than 40%, it is unlikely that <str<strong>on</strong>g>the</str<strong>on</strong>g> success <str<strong>on</strong>g>of</str<strong>on</strong>g><br />
E. litoralis can be attributed solely to its reproductive activities.<br />
Enoplolaimus litoralis is assumed to be a predacious nematode,<br />
because <str<strong>on</strong>g>of</str<strong>on</strong>g> its large buccal cavity and associated teeth and mandibles,<br />
and <str<strong>on</strong>g>the</str<strong>on</strong>g>refore may be less dependent <strong>on</strong> <str<strong>on</strong>g>the</str<strong>on</strong>g> biochemistry <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
sediment, compared to detritivorous or microvorous species. Hence,<br />
it seems to be less hampered in col<strong>on</strong>ising <str<strong>on</strong>g>the</str<strong>on</strong>g> available sediments<br />
than are o<str<strong>on</strong>g>the</str<strong>on</strong>g>r species. <str<strong>on</strong>g>The</str<strong>on</strong>g>refore, <str<strong>on</strong>g>the</str<strong>on</strong>g> col<strong>on</strong>isati<strong>on</strong> success <str<strong>on</strong>g>of</str<strong>on</strong>g><br />
E. litoralis likely results from <str<strong>on</strong>g>the</str<strong>on</strong>g> combinati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> its high mobility and<br />
feeding activity. This species may also be resp<strong>on</strong>sible for <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
reducti<strong>on</strong> in diversity, by preying <strong>on</strong> o<str<strong>on</strong>g>the</str<strong>on</strong>g>r n<strong>on</strong>-predatory species<br />
(i.e., a top-down <str<strong>on</strong>g>effect</str<strong>on</strong>g>). Top-down <str<strong>on</strong>g>effect</str<strong>on</strong>g>s <str<strong>on</strong>g>of</str<strong>on</strong>g> predatory nematodes<br />
have been shown to c<strong>on</strong>trol <str<strong>on</strong>g>the</str<strong>on</strong>g> density and species compositi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g><br />
nematode communities (Moens et al., 2000; Steyaert et al., 2001;<br />
Gallucci et al., 2005; Dos Santos and Moens, 2011).<br />
From day 14 <strong>on</strong>wards, <str<strong>on</strong>g>the</str<strong>on</strong>g>re was a slight increase in richness and<br />
diversity in both treatments. This could indicate better sediment<br />
c<strong>on</strong>diti<strong>on</strong>s developed through col<strong>on</strong>isati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> sediment by<br />
microbes and diatoms from <str<strong>on</strong>g>the</str<strong>on</strong>g> water column. Although <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
recovery <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> initial compositi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> bacterial community may<br />
occur 25 days after defaunati<strong>on</strong>, <str<strong>on</strong>g>the</str<strong>on</strong>g> highest density <str<strong>on</strong>g>of</str<strong>on</strong>g> microbial<br />
cells may be found <strong>on</strong> around <str<strong>on</strong>g>the</str<strong>on</strong>g> tenth day <str<strong>on</strong>g>of</str<strong>on</strong>g> a microbial col<strong>on</strong>isati<strong>on</strong><br />
process (Stocum and Plante, 2006).<br />
Table 3<br />
Output <str<strong>on</strong>g>of</str<strong>on</strong>g> two-way crossed SIMPER analysis showing <str<strong>on</strong>g>the</str<strong>on</strong>g> top 50% typical species for<br />
each treatment. Overall similarity <str<strong>on</strong>g>of</str<strong>on</strong>g> each treatment is shown between brackets.<br />
Abbreviati<strong>on</strong>s as used in Table 1.<br />
Species AQ (72%) IC (69%) AT (63%) ST (63%)<br />
Sigmophoranema rufum 10 9<br />
Enoplolaimus litoralis 8 8 32 20<br />
Mesacanthi<strong>on</strong> sp. 1 7 8 10 10<br />
Oncholaimellus calvadosicus 7 6 7<br />
Ascolaimus el<strong>on</strong>gatus 6 10 9<br />
Chromadora axi 6 6<br />
Metadesmolaimus sp. 1 6 6<br />
Neochromadora munita 6 5<br />
Dapt<strong>on</strong>ema normandicus 5 10<br />
T.F. Maria et al. / Estuarine, Coastal and Shelf Science 94 (2011) 272e280 277<br />
4.3. Community shifts in <str<strong>on</strong>g>the</str<strong>on</strong>g> presence and absence <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>Scolelepis</str<strong>on</strong>g><br />
<str<strong>on</strong>g>squamata</str<strong>on</strong>g> after 2 weeks <str<strong>on</strong>g>of</str<strong>on</strong>g> col<strong>on</strong>isati<strong>on</strong><br />
Significant differences in nematode diversity and <str<strong>on</strong>g>the</str<strong>on</strong>g> density <str<strong>on</strong>g>of</str<strong>on</strong>g><br />
Enoplolaimus litoralis between <str<strong>on</strong>g>the</str<strong>on</strong>g> azoic treatment and <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>Scolelepis</str<strong>on</strong>g><br />
treatment occurred at day 14, and indeed PERMANOVA indicated<br />
<str<strong>on</strong>g>the</str<strong>on</strong>g> lowest n<strong>on</strong>-significant difference in nematode<br />
community compositi<strong>on</strong> between <str<strong>on</strong>g>the</str<strong>on</strong>g>se treatments at this time.<br />
<str<strong>on</strong>g>The</str<strong>on</strong>g>se results indicate that <str<strong>on</strong>g>the</str<strong>on</strong>g> presence <str<strong>on</strong>g>of</str<strong>on</strong>g> S. <str<strong>on</strong>g>squamata</str<strong>on</strong>g> affects <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
col<strong>on</strong>isati<strong>on</strong>, rejecting our sec<strong>on</strong>d null hypo<str<strong>on</strong>g>the</str<strong>on</strong>g>sis (H2).<br />
In <str<strong>on</strong>g>the</str<strong>on</strong>g> presence <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>polychaete</str<strong>on</strong>g>, Enoplolaimus litoralis<br />
appeared in lower densities, from day 14 compared to when <str<strong>on</strong>g>Scolelepis</str<strong>on</strong>g><br />
<str<strong>on</strong>g>squamata</str<strong>on</strong>g> was absent. <str<strong>on</strong>g>The</str<strong>on</strong>g> <str<strong>on</strong>g>polychaete</str<strong>on</strong>g> seem to reduce <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
top-down <str<strong>on</strong>g>effect</str<strong>on</strong>g> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> predatory nematode, favouring earlier<br />
establishment <str<strong>on</strong>g>of</str<strong>on</strong>g> additi<strong>on</strong>al n<strong>on</strong>-predatory species (i.e., Dapt<strong>on</strong>ema<br />
normandicus), which in turn led to an increase in diversity (H 0 ). In<br />
additi<strong>on</strong> to delaying/inhibiting <str<strong>on</strong>g>the</str<strong>on</strong>g> col<strong>on</strong>isati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> E. litoralis, <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
<str<strong>on</strong>g>polychaete</str<strong>on</strong>g> may also help in enhancing <str<strong>on</strong>g>the</str<strong>on</strong>g> envir<strong>on</strong>mental c<strong>on</strong>diti<strong>on</strong>s,<br />
again c<strong>on</strong>tributing to <str<strong>on</strong>g>the</str<strong>on</strong>g> higher nematode diversity in <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
<str<strong>on</strong>g>Scolelepis</str<strong>on</strong>g> treatment. Better envir<strong>on</strong>mental c<strong>on</strong>diti<strong>on</strong>s can result<br />
from <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>polychaete</str<strong>on</strong>g>’s activities, which change <str<strong>on</strong>g>the</str<strong>on</strong>g> sediment<br />
chemistry through bioturbati<strong>on</strong> and/or increase microbial metabolism<br />
by depositi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> pseud<str<strong>on</strong>g>of</str<strong>on</strong>g>aeces (Dauer, 1983; Hartmann-<br />
Schröeder, 1996; Pardo and Amaral, 2004; Van Hoey et al., 2004).<br />
On <str<strong>on</strong>g>the</str<strong>on</strong>g> o<str<strong>on</strong>g>the</str<strong>on</strong>g>r hand, <str<strong>on</strong>g>polychaete</str<strong>on</strong>g> bioturbati<strong>on</strong> stimulates organicmatter<br />
mineralizati<strong>on</strong> via microbial degradati<strong>on</strong>, by increasing<br />
<str<strong>on</strong>g>the</str<strong>on</strong>g> oxygen supply from its burrow c<strong>on</strong>structi<strong>on</strong> and irrigati<strong>on</strong><br />
(Mermillod-Bl<strong>on</strong>din et al., 2004; Papaspyrou et al., 2007;<br />
Timmermann et al., 2008; Braeckman et al., 2010). <str<strong>on</strong>g>The</str<strong>on</strong>g> individuals<br />
<str<strong>on</strong>g>of</str<strong>on</strong>g> S. <str<strong>on</strong>g>squamata</str<strong>on</strong>g> built temporary vertical tubes and occasi<strong>on</strong>ally<br />
burrow to a depth <str<strong>on</strong>g>of</str<strong>on</strong>g> 40 cm below <str<strong>on</strong>g>the</str<strong>on</strong>g> sediment surface (Hartmann-<br />
Schröeder, 1996; Van Hoey et al., 2004). Indeed, during <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
sampling times, some burrows were found at <str<strong>on</strong>g>the</str<strong>on</strong>g> bottom <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
cores, indicating activity <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>polychaete</str<strong>on</strong>g>.<br />
4.4. Successi<strong>on</strong> patterns in c<strong>on</strong>trasting envir<strong>on</strong>ments<br />
Three generalized models <str<strong>on</strong>g>of</str<strong>on</strong>g> community recovery were<br />
proposed by C<strong>on</strong>nell and Slatyer (1977), based <strong>on</strong> interacti<strong>on</strong>s<br />
between pi<strong>on</strong>eering species and later col<strong>on</strong>ists: 1- Facilitati<strong>on</strong><br />
model: early col<strong>on</strong>ists (i.e., pi<strong>on</strong>eering) can promote <str<strong>on</strong>g>the</str<strong>on</strong>g> establishment<br />
<str<strong>on</strong>g>of</str<strong>on</strong>g> later col<strong>on</strong>ists; 2- Inhibiti<strong>on</strong> model: early col<strong>on</strong>ists<br />
reduce <str<strong>on</strong>g>the</str<strong>on</strong>g> establishment <str<strong>on</strong>g>of</str<strong>on</strong>g> later col<strong>on</strong>ists; and 3- Tolerance<br />
model: early col<strong>on</strong>ists have little or no <str<strong>on</strong>g>effect</str<strong>on</strong>g> <strong>on</strong> <str<strong>on</strong>g>the</str<strong>on</strong>g> establishment<br />
<str<strong>on</strong>g>of</str<strong>on</strong>g> later col<strong>on</strong>ists. In mudflats, an envir<strong>on</strong>ment with low hydrodynamic<br />
stress, very fine sediment and naturally high abundance <str<strong>on</strong>g>of</str<strong>on</strong>g><br />
microphytobenthos (Reise, 1985), opportunistic epistrate-feeding<br />
nematode species dominate in <str<strong>on</strong>g>the</str<strong>on</strong>g> absence <str<strong>on</strong>g>of</str<strong>on</strong>g> macr<str<strong>on</strong>g>of</str<strong>on</strong>g>auna. This is<br />
explained by <str<strong>on</strong>g>the</str<strong>on</strong>g> high availability <str<strong>on</strong>g>of</str<strong>on</strong>g> microphytobenthos in <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
absence <str<strong>on</strong>g>of</str<strong>on</strong>g> grazing macr<str<strong>on</strong>g>of</str<strong>on</strong>g>auna (Van Colen et al., 2009). In this case,<br />
early-col<strong>on</strong>ising nematodes c<strong>on</strong>tributed to a delay in <str<strong>on</strong>g>the</str<strong>on</strong>g> nematode<br />
community recovery, and for that reas<strong>on</strong> an inhibiti<strong>on</strong> model was<br />
recognized. But in plots where macr<str<strong>on</strong>g>of</str<strong>on</strong>g>auna was still present, epistrate<br />
nematodes did not reach high densities, suggesting that <str<strong>on</strong>g>the</str<strong>on</strong>g>ir<br />
low densities provide stability for <str<strong>on</strong>g>the</str<strong>on</strong>g> nematode community (Van<br />
Colen et al., 2009).<br />
In sandy beaches, an envir<strong>on</strong>ment that sharply diverges from<br />
a mudflat because <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> str<strong>on</strong>ger hydrodynamics, coarse sediments<br />
and low abundance <str<strong>on</strong>g>of</str<strong>on</strong>g> microphytobenthos (McLachlan and<br />
Brown, 2006), <str<strong>on</strong>g>the</str<strong>on</strong>g> presence <str<strong>on</strong>g>of</str<strong>on</strong>g> macrobenthos seems to lead to<br />
a delay/inhibiti<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> establishment <str<strong>on</strong>g>of</str<strong>on</strong>g> high densities <str<strong>on</strong>g>of</str<strong>on</strong>g> opportunistic<br />
species, here represented by Enoplolaimus litoralis. <str<strong>on</strong>g>The</str<strong>on</strong>g> low<br />
density <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>dominant</str<strong>on</strong>g> predatory nematode species favours <str<strong>on</strong>g>the</str<strong>on</strong>g><br />
earlier establishment <str<strong>on</strong>g>of</str<strong>on</strong>g> sec<strong>on</strong>dary species (Vanaverbeke et al.,<br />
2007). In our experiment, in <str<strong>on</strong>g>the</str<strong>on</strong>g> absence <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>polychaete</str<strong>on</strong>g>, <str<strong>on</strong>g>the</str<strong>on</strong>g>














