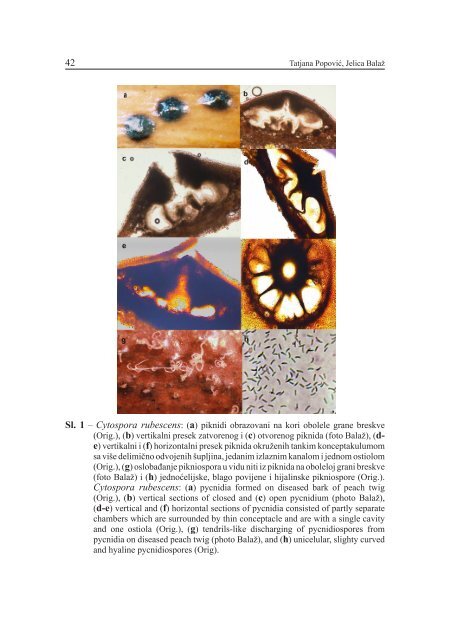
42 Tatjana Popović, Jelica Balaž Sl. 1 – Cytospora rubescens: (a) piknidi obrazovani na kori obolele grane breskve (Orig.), (b) vertikalni presek zatvorenog i (c) otvorenog piknida (foto Balaž), (de) vertikalni i (f) horizontalni presek piknida okruženih tankim konceptakulumom sa više delimično odvojenih šupljina, jedanim izlaznim kanalom i jednom ostiolom (Orig.), (g) oslobađanje pikniospora u vidu niti iz piknida na oboleloj grani breskve (foto Balaž) i (h) jednoćelijske, blago povijene i hijalinske pikniospore (Orig.). Cytospora rubescens: (a) pycnidia formed on diseased bark of peach twig (Orig.), (b) vertical sections of closed and (c) open pycnidium (photo Balaž), (d-e) vertical and (f) horizontal sections of pycnidia consisted of partly separate chambers which are surrounded by thin conceptacle and are with a single cavity and one ostiola (Orig.), (g) tendrils-like discharging of pycnidiospores from pycnidia on diseased peach twig (photo Balaž), and (h) unicelular, slighty curved and hyaline pycnidiospores (Orig).
Anatomsko-histološke karakteristike Leucostoma cincta parazita breskve 43 Na presecima piknidijalnine strome uočava se veći broj odvojenih ili delimično odvojenih komora povezanih zajedničkim izlaznim otvorom i jednom ostiolom za oslobađanje pikniospore (sl.1)., što je tipično za vrstu C. rubescens (Hayova et Minter, 1998). Prema Gvritishvili (1982) konceptakulum je kod vrste C. rubescens debljine do 150 ìm ili ponekad i odsustvuje, dok je kod slične vrste C. leucostoma debljine 50-200 ìm i tamno-mrke do crne boje. Piknidiospore. Oslobađanje pikniospora iz piknida je u vidu tankih niti, oranž do crvene boje (sl.7) na inokulisanim reznicama i granama breskve, ali je oslobađanje u vidu kapi ili končića retko na KDA. Ovakav način izlučivanja pikniospora je karakteristika vrste (Vajna, 1981; Arsenijević i Balaž, 1982; Gvritishvili, 1982; Biggs, 1997; Hayova et Minter, 1998). Prema navodima Rozsnyay (1977), crvena masa pikniospora se oslobađa i širi preko površine u vidu kapi ukoliko je vreme vlažno, dok je po suvom vremenu izbacivanje spora u vidu niti. Proces pražnjenja piknida odvijao se po hladnom i vlažnom vremenu tokom naših ogleda. Izbačene pikniospore se često mogu videti posle kiše (Rozsnyay, 1977; Arsenijević i Balaž, 1982; Biggs, 1997; Ivanović i Ivanović, 2001; Tisserat, 2003). S druge strane, Luepschen et Rohrbach (1969) navode da se oslobađanje pikniospora najviše odvija tokom leta, što nije u saglasnosti sa našim zapažanjima. Formiranje pikniospore je uglavnom tokom proleća i jeseni, a ređe zimi (Ivanović i Ivanović, 2001), mada pikniospore kao osnovni izvor zaraze mogu biti prisutne u voćnjaku praktično tokom cele godine (Rozsnyay, 1977; Arsenijević i Balaž, 1982). Pikniospore su jednoćelične, blago povijene, tankih zidova, hijalinske (sl.1), što odgovara opisima drugih autora (Hayova et Minter, 1998; Gvritishvili, 1982; Biggs, 1997). Dimenzije pikniospora iz piknida obrazovanih na inokulisanim reznicama su 5,0 – (6,5) – 8,5 x 1,5 – (1,5) – 2,5 µm. Ove vrednosti su u granicama citiranih veličina piknidiospora: u kulturi 5,0 – (5,5)– 7,5 x 1,2– (1,4) – 1,7 µm, odnosno 5,0 – (6,9) – 8,7 x 1,2 – (1,5) – 1,7 µm na obolelom tkivu (Arsenijević, 1973), 4-6,5 x 1-1,5 µm (Vajna, 1981), 5-13 x 1,5-3 µm (Gvritishvili, 1982), 5-10 x 1-2 ìm (Biggs, 1997) i 5-8 (-13) x 1-3 µm (Hayova et Minter, 1998). Obrazovanje peritecija. Višegodišnjim ispitivanjima (1997.-2002.) obolelih grana breskve nije konstatovano formiranje savršenog stadijuma gljive. Takođe, peritecijske stromate nisu obrazovane in-vitro na podlozi. Za obrazovanje peritecija potreban je dug period, često 2-3 godine posle formiranja piknidske strome (Biggs, 1997). Izgleda da telemorf nema veći značaj u biologiji patogena, s obzirom da njegovo odsustvo u ciklusu razvoja gljive navode i drugi istraživači (Kable et al., 1967; Rozsnyay et al., 1972, loc. cit. Rozsnyay, 1977; Rozsnyay et Klement,