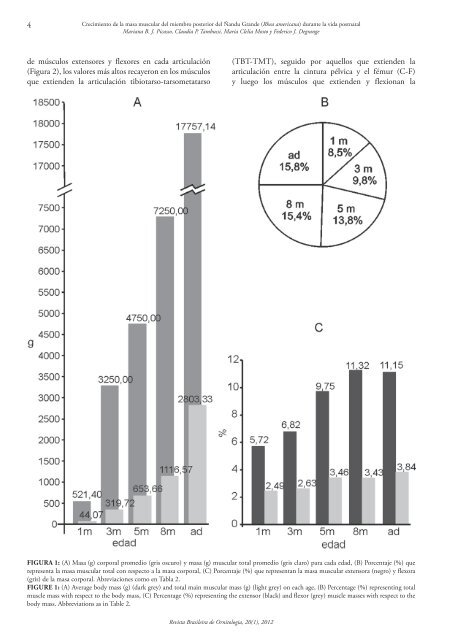
4Crecimiento <strong>de</strong> la masa muscular <strong>de</strong>l miembro posterior <strong>de</strong>l Ñandu Gran<strong>de</strong> (Rhea americana) durante la vida postnatalMariana B. J. Picasso, Claudia P. Tambussi, María Clelia Mosto y Fe<strong>de</strong>rico J. Degrange<strong>de</strong> músculos extensores y flexores en cada articulación(Figura 2), los valores más altos recayeron en los músculosque extien<strong>de</strong>n la articulación tibiotarso-tarsometatarso(TBT‐TMT), seguido por aquellos que extien<strong>de</strong>n laarticulación entre la cintura pélvica y el fémur (C‐F)y luego los músculos que extien<strong>de</strong>n y flexionan laFigura 1: (A) Masa (g) corporal promedio (gris oscuro) y masa (g) muscular total promedio (gris claro) para cada edad, (B) Porcentaje (%) querepresenta la masa muscular total con respecto a la masa corporal, (C) Porcentaje (%) que representan la masa muscular extensora (negro) y flexora(gris) <strong>de</strong> la masa corporal. Abreviaciones como en Tabla 2.Figure 1: (A) Average body mass (g) (dark grey) and total main muscular mass (g) (light grey) on each age, (B) Percentage (%) representing totalmuscle mass with respect to the body mass, (C) Percentage (%) representing the extensor (black) and flexor (grey) muscle masses with respect to thebody mass. Abbreviations as in Table 2.Revista <strong>Brasileira</strong> <strong>de</strong> <strong>Ornitologia</strong>, <strong>20</strong>(1), <strong>20</strong>12
Crecimiento <strong>de</strong> la masa muscular <strong>de</strong>l miembro posterior <strong>de</strong>l Ñandu Gran<strong>de</strong> (Rhea americana) durante la vida postnatalMariana B. J. Picasso, Claudia P. Tambussi, María Clelia Mosto y Fe<strong>de</strong>rico J. Degrange5articulación entre el fémur y el tibiotarso (F‐TBT).Sólo los músculos que flexionan los dígitos superaron aaquellos que los extien<strong>de</strong>n.DiscusiónMasa muscular total y masa corporalEn las aves, la simple relación entre la masa muscular<strong>de</strong> los miembros y la masa corporal brinda informaciónsobre el <strong>de</strong>sarrollo y la actividad que la musculatura realizadurante la locomoción (Hartman 1961). Los valoreshallados en los adultos <strong>de</strong>l Ñandú Gran<strong>de</strong> son similaresa los encontrados en otras Ratites: el Avestruz (Struthiocamelus) (Smith et al. <strong>20</strong>06) y en en el Emú (Dromaiusnovaehollandiae) (Patak y Baldwin 1993), don<strong>de</strong> la masamuscular <strong>de</strong> ambos miembros posteriores representó el33% y el 25% <strong>de</strong> la masa corporal respectivamente. Estosvalores son altos al compararlos con aves que presentanestilos locomotores no cursoriales como los Hirundinidaey los Trochilidae cuyo porcentaje es <strong>de</strong>l 1% o en losTinamidae y los Cracidae con valores <strong>de</strong>l 19% (Hartman1961). La alta proporción <strong>de</strong> la musculatura <strong>de</strong>l miembroposterior <strong>de</strong> Rhea americana es indicativa <strong>de</strong>l amplio grado<strong>de</strong> <strong>de</strong>sarrollo y actividad que el miembro <strong>de</strong>sempeñadurante la locomoción (Picasso <strong>20</strong>10b). Esta importanciaqueda <strong>de</strong> manifiesto <strong>de</strong>s<strong>de</strong> las etapas más tempranas <strong>de</strong>la vida postnatal. Lamentablemente no hay información<strong>de</strong> este tipo para el resto <strong>de</strong> las eda<strong>de</strong>s que no sea la <strong>de</strong>lFigura 2: Porcentaje (%) <strong>de</strong> la masa corporal que representan losmúsculos extensores (columna <strong>de</strong> la izquierda) y flexores (columna<strong>de</strong> la <strong>de</strong>recha) en cada articulación. CF (articulación cintura pélvica‐-fémur), F‐TBT (articulación fémur-tibiotarso), TBT‐TMT (articulacióntibiotarso-tarsometatarso).Figure 2: Percentage (%) of body mass represented by the extensor(left column) and flexors (right column) muscles at each joint.CF (pelvic girdle-femur joint), F‐TBT (femur-tibiotarsus joint),TBT‐TMT (tibiotarsus-tarsometatarsus joint).adulto en aves con otros estilos locomotores que permitanla comparación con R. americana.Los músculos extensores y flexoresEl predominio <strong>de</strong> la masa extensora, evi<strong>de</strong>nciadoen todas las eda<strong>de</strong>s, pue<strong>de</strong> estar asociado a dos factores.Por un lado, en la locomoción terrestre los músculosextensores están comprometidos en el sostén <strong>de</strong>l cuerpo yen su aceleración y <strong>de</strong>saceleración (Gatesy 1999a) <strong>de</strong>bidoa que son los movimientos <strong>de</strong> extensión los que actúancuando el miembro es apoyado en el suelo (fase <strong>de</strong> apoyo)durante la locomoción. Esto genera un mayor stressmuscular contribuyendo, entonces, al mayor <strong>de</strong>sarrollo <strong>de</strong>estos. Por otro lado, el mayor <strong>de</strong>sarrollo <strong>de</strong> la musculaturaextensora es una característica <strong>de</strong> animales corredorescomo el caballo (Payne et al. <strong>20</strong>05) y <strong>de</strong> las gran<strong>de</strong>sRatites como el Avestruz (Struthio camelus) (Smith et al.<strong>20</strong>06) y el Emú (Dromaius novaehollandiae) (Hutchinson<strong>20</strong>04). Al comparar estos datos con aquellos disponiblespara aves <strong>de</strong> locomoción terrestre pero no especializadasen la carrera como el Pavo Común (Meleagris gallopavo,Galliformes) y la Martineta Común (Eudromia elegans,Tinamiformes), el porcentaje <strong>de</strong> masa muscular extensorafue menor (5,4% en Meleagris, Roberts et al. 1998 y 4%en Eudromia, Hutchinson <strong>20</strong>04) a los registrados aquípara el Ñandú Gran<strong>de</strong>.Los músculos extensores <strong>de</strong> la articulaciónTBT‐TMT predominaron sobre los extensores <strong>de</strong> lasarticulaciones restantes en todas las eda<strong>de</strong>s estudiadas.Los movimientos <strong>de</strong> extensión <strong>de</strong> esta articulación sonimportantes ya que alargan el miembro y permitenalcanzar mayor velocidad y aceleración (Smith et al.<strong>20</strong>06), un rasgo esencial en animales corredores. Enconsecuencia, el <strong>de</strong>sarrollo marcado <strong>de</strong> los extensores <strong>de</strong>lTBT‐TMT parece estar estrechamente vinculado al tipo<strong>de</strong> locomoción cursorial. Los extensores <strong>de</strong> la articulaciónC‐F siguieron en importancia a aquellos <strong>de</strong>l TBT‐TMT.Estos músculos poseen roles muy importantes en lalocomoción terrestre y, en especial, en aves corredoras,<strong>de</strong>bido a que su acción brinda la fuerza necesaria parasoportar la masa corporal y minimizar los movimientos<strong>de</strong> balanceo <strong>de</strong>l cuerpo durante la carrera, cuando cadamiembro alternadamente <strong>de</strong>be soportar la masa corporal(Berger 1952, Sasaki y Neptune <strong>20</strong>06). A<strong>de</strong>más, suacción extensora colabora, junto con los extensores <strong>de</strong>lTBT‐TMT, con el aumento <strong>de</strong> la velocidad durante lacarrera (Gatesy y Biewener 1991, Reilly <strong>20</strong>00).El tercer grupo en importancia en cuanto a losproporciones respecto a la masa corporal es el <strong>de</strong> losextensores <strong>de</strong> la articulación F‐TBT (M. femorotibialis),alcanzando valores semejantes a los que Hutchinson(<strong>20</strong>04) brinda para un ave caminadora como la MartinetaComún (Eudromia elegans). Por lo tanto, el <strong>de</strong>sarrollo <strong>de</strong>Revista <strong>Brasileira</strong> <strong>de</strong> <strong>Ornitologia</strong>, <strong>20</strong>(1), <strong>20</strong>12