Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Χαρακτηριστική ιδιότητα και λειτουργία των ενζύμων, είναι η<br />
κατάλυση των χημικών αντιδράσεων.<br />
• Μελέτη της καταλυτικής δράσης, πρέπει να βασίζεται στον<br />
ποσοτικό προσδιορισμό της ταχύτητας της χημικής<br />
αντίδρασης που καταλύεται.<br />
• Από τη μελέτη της μεταβολής της ταχύτητας, καταλήγουμε<br />
σε συμπεράσματα χρήσιμα για τη δομή και για το μηχανισμό<br />
δράσης των ενζύμων ( κυρίως όταν το ένζυμο είναι υψηλής<br />
καθαρότητας).<br />
•Η μελέτη της ενζυμικής κινητικής είναι σημαντική και για<br />
πρακτικούς λόγους:<br />
• Ο ορισμός ικανοποιητικών μονάδων (Units) απαιτεί να<br />
γνωρίζουμε<br />
• τις βέλτιστες συνθήκες (pΗ, θερμοκρασία κ.λπ.) για τη<br />
δράση του ενζύμου, καθώς και<br />
• την επίδραση των διαφόρων παραγόντων<br />
(αναστολείς, ενεργοποιητές κ.λπ.) σ’ αυτήν.
• Η γνώση της ενζυμικής κινητικής βοηθάει:<br />
• στην κατανόηση των βιολογικών φαινομένων τα οποία,<br />
είναι πολύ ευαίσθητα σε αλλαγές θερμοκρασίας, pΗ κ.λπ.<br />
• στην επίλυση του ερωτήματος “με ποιο τρόπο δρουν τα<br />
ένζυμα μέσα στο κύτταρο”.<br />
• Για όλους τους παραπάνω λόγους, η ενζυμική κινητική ή<br />
κινητική ενζυμικών αντιδράσεων, έχει εξελιχθεί σε ένα<br />
ιδιαίτερο επιστημονικό κλάδο.<br />
• Στην ενζυμική κινητική πρωταρχικό ρόλο παίζει η μελέτη<br />
της ταχύτητας (v) της ενζυμικής αντίδρασης σε συνάρτηση με<br />
τους διάφορους παράγοντες που επιδρούν σ’ αυτήν,<br />
• με επίκεντρο τη μελέτη της σχέσης μεταξύ της ταχύτητας<br />
της ενζυμικής αντίδρασης και της συγκέντρωσης<br />
• του ενζύμου (Ε) και<br />
• του υποστρώματος (S).
ΜΕΤΡΗΣΗ ΤΗΣ ΚΑΤΑΛΥΤΙΚΗΣ ΔΡΑΣΗΣ ΤΩΝ<br />
ΕΝΖΥΜΩΝ<br />
• Μερικά μόνο ένζυμα μπορούν να προσδιοριστούν με<br />
άμεσες φασματομετρικές μεθόδους.<br />
• Η παρουσία των περισσοτέρων ενζύμων πιστοποιείται<br />
από τη θετική έκβαση των συγκεκριμένων αντιδράσεων που<br />
καταλύουν<br />
• Το ποσό των ενζύμων υπολογίζεται από την ταχύτητα της<br />
αντίδρασης.<br />
• Η μέτρηση λοιπόν της ταχύτητας της αντίδρασης, είναι το<br />
πιο σημαντικό κομμάτι των τεχνικών που χρησιμοποιούνται<br />
στη μελέτη των ενζύμων.
• Η μορφή της καμπύλης της αλλοίωσης του υποστρώματος<br />
ήτηςπαραγωγής του προϊόντος συναρτήσει του βαθμού<br />
προόδου της αντίδρασης, στις περισσότερες ενζυμικές<br />
αντιδράσεις, είναι της γενικής μορφής του σχήματος (η<br />
ταχύτητα μειώνεται συναρτήσει του χρόνου).<br />
•Το φαινόμενο αυτό μπορεί να οφείλεται σε πολλούς λόγους:<br />
• π.χ. τα προϊόντα της αντίδρασης αναστέλλουν το ένζυμο<br />
• ο βαθμός κορεσμού του<br />
ενζύμου από το υπόστρωμα<br />
μειώνεται πιθανώς λόγω μείωσης<br />
της συγκέντρωσης του υποστρώματος<br />
συναρτήσει του βαθμού<br />
προόδου της αντίδρασης,<br />
• η αντίστροφη πορεία της αντίδρασης<br />
γίνεται σε μεγαλύτερο<br />
βαθμό λόγω της αύξησης της<br />
συγκέντρωσης των προϊόντων<br />
κ.λπ.
• Για να αποφευχθούν οι παραπάνω λόγοι, υιοθετήθηκε η<br />
μέτρηση της αρχικής ταχύτητας, κατά την οποία οι<br />
παραπάνω λόγοι δεν έχουν τη χρονική δυνατότητα να<br />
παρέμβουν.<br />
• Κατά τη μελέτη των ενζύμων, είναι απόλυτα αναγκαίο να<br />
προσδιορίζεται ηεπίδρασηστηναρχικήταχύτηταενός<br />
παράγοντα (pΗ, θερμοκρασία, συγκέντρωση ενζύμου κ.λπ.)<br />
κάθε φορά.<br />
• Για τη μέτρηση της αρχικής ταχύτητας, επιλέγεται συνήθως<br />
ένας τέτοιος χρόνος, ώστε η αλλοίωση του υποστρώματος,<br />
να είναι κάτω από το 20% της συνολικής.<br />
• Μέχρι αυτού του ορίου, οι καμπύλες της ταχύτητας, σε<br />
συνάρτηση με το μεταβαλλόμενο παράγοντα είναι συνήθως<br />
ευθείες γραμμές.
•Για να ορισθεί η αρχική ταχύτητα, γίνεται μία σειρά πειραμάτων,<br />
για να καθορισθούν τα όρια της γραμμικότητας, ώστε<br />
• η παραγωγή του προϊόντος να είναι μέγιστη και<br />
• οι καμπύλες της ταχύτητας συναρτήσει του χρόνου και<br />
της συγκέντρωσης του ενζύμου [ v = f(t) και v = f([Ε])], να<br />
είναι ευθείες.<br />
•Καθορισμός αρχικής<br />
ταχύτητας:<br />
•Προσδιορίζεται<br />
το Ρ = f (t), όταν<br />
όλοι οι άλλοι παράμετροι,<br />
είναι σταθερές<br />
(π.χ.[S], pΗ,<br />
T κ.λπ.) εκτός [E].
• Κατασκευάζεται υπολογιστικά [από το διάγραμμα<br />
Ρ = f (t)], το διάγραμμα v = f([Ε]).<br />
•Το διάγραμμα v = f([Ε]) για χρόνο t 0 είναι ευθεία γραμμή,<br />
δηλαδή η ταχύτητα είναι ανάλογη της συγκέντρωσης του<br />
ενζύμου και αντιπροσωπεύει την αρχική ταχύτητα.<br />
• Στο διάγραμμα v = f([Ε]) για χρόνο t 1 , υπάρχει ένα ευθύγραμμο<br />
τμήμα, που<br />
συμπίπτει με εκείνο<br />
του χρόνου t 0 , ενώ<br />
στο διάγραμμα<br />
v = f([Ε]), που αντιστοιχεί<br />
σε χρόνο t 2 ,<br />
ηταχύτηταδενείναι<br />
ανάλογη της συγκέντρωσης<br />
του<br />
ενζύμου.
• Από τα παραπάνω φαίνεται ότι η καμπύλη v = f([Ε]) είναι<br />
ευθεία, μόνο όταν χρησιμοποιούνται αρχικές ταχύτητες.<br />
• Επομένως, γιανααναφερόμαστεσεαρχικήταχύτητα, θα<br />
πρέπει να χρησιμοποιείται<br />
συγκέντρωση<br />
ενζύμου ίση με<br />
[Ε 1 ] και χρόνος t 1 .<br />
•Τότε θα ικανοποιούνται<br />
οι παραπάνω<br />
απαιτήσεις,<br />
δηλαδή η παραγωγή<br />
του προϊόντος<br />
να είναι<br />
μέγιστη, αλλά και<br />
οι καμπύλες v = f(t)<br />
και v = f([Ε]), να<br />
είναι ευθείες.
• Υπάρχουν όμως περιπτώσεις, όπου το διάγραμμα<br />
προόδου της αντίδρασης δεν έχει τη μορφή του σχήματος:<br />
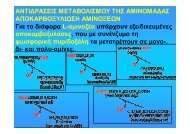
• Η οξειδάση των D-αμινοξέων<br />
απαιτεί για τη δράση της, την<br />
παρουσία φλαβινοσυνενζύμων,<br />
(στην πραγματικότητα<br />
είναι προσθετικές<br />
ομάδες) που συνδέονται με<br />
το ένζυμο με αργό ρυθμό.<br />
• Αυτό έχει σαν αποτέλεσμα<br />
να αυξάνεται δραστικά το<br />
ποσό του ενεργού ενζύμου<br />
μετά την πάροδο κάποιου<br />
χρονικού διαστήματος, από τη στιγμή της ανάμιξης των<br />
αντιδρώντων.<br />
• Το πρόβλημα αυτό θα μπορούσε να αντιμετωπιστεί με<br />
προεπώαση ενζύμου και συνενζύμου (ή προσθετικής<br />
ομάδας), πριν από την προσθήκη του υποστρώματος.
• Όταν αναφέρεται η λέξη ταχύτητα θα εννοείται πάντα η<br />
αρχική ταχύτητα εκτός αν διευκρινίζεται κάτι διαφορετικό.<br />
• Η παρακολούθηση του βαθμού προόδου μιας ενζυμικής<br />
αντίδρασης μπορεί να γίνει:<br />
• με τη λήψη δείγματος σε διάφορες διαδοχικές χρονικές<br />
στιγμές από το ίδιο μίγμα αντίδρασης (sampling methods)<br />
• με τη χρήση διαφορετικών μιγμάτων αντίδρασης που<br />
έχουν ίδια σύσταση και τα οποία επωάζονται σε<br />
διαφορετικούς χρόνους<br />
(continuous methods),<br />
μέθοδος που έχει<br />
μεγαλύτερη εφαρμογή.
Ποσοτικός προσδιορισμός των ενζύμων<br />
• Τα ένζυμα είναι πρωτεΐνες και συνυπάρχουν με άλλες<br />
πρωτεϊνες και ενώσεις, καθιστώντας το χημικό<br />
προσδιορισμό τους σχεδόν αδύνατο.<br />
• Η ποσότητα του ενζύμου, υπολογίζεται με προσδιορισμό<br />
της καταλυτικής της δράσης, η οποία εκφράζεται ποσοτικά<br />
με τους παρακάτω όρους (σύμφωνα με τη Commision on<br />
Εnzymes).<br />
• Μονάδες ενζυμικής δραστικότητας<br />
[U = μmol[S]/min]<br />
1) Ειδική δραστικότητα [ U/mg πρωτεΐνης]<br />
2) Μοριακή δραστικότητα [ U/μmol ενζύμου ]<br />
3) Δραστικότητα<br />
καταλυτικού κέντρου<br />
[Αλλοιωμένα μόρια S / min<br />
και προσθετική ομάδακαταλυτικό<br />
κέντρο]
• Μονάδα ενζυμικής δραστικότητας (1U: Ιnternational Unit):<br />
Το ποσό του E που αλλοιώνει 1 μmol S (ή παράγει 1 μmol P)<br />
ανά 1 min, σε καθορισμένες συνθήκες.<br />
•Όταν επηρεάζονται περισσότεροι από ένα δεσμοί σε κάθε<br />
μόριο S: Το ποσό του Ε που καταλύει τη μετατροπή 1<br />
μequivalent της ομάδας που επηρεάζεται ανά 1 min.<br />
• Στις καθορισμένες συνθήκες περιλαμβάνεται<br />
• η θερμοκρασία (αρχικά είχε ορισθεί 25 0 C μετά 30 0 C, 37 0 C<br />
ενώ σήμερα χρησιμοποιείται η θερμοκρασία όπως<br />
ορίζεται για κάθε περίπτωση)<br />
• το pΗ και<br />
• η συγκέντρωση του S.<br />
(Για τα δύο τελευταία χρησιμοποιούνται συνήθως οι<br />
βέλτιστες συνθήκες για το κάθε ένζυμο).<br />
• Όταν αντιδρούν δύο ίδια μόρια S μεταξύ τους: Το ποσό του<br />
E που καταλύει την αλλοίωση 2 μmoles S σε 1 min.<br />
( μονάδα απόλυτη που επιτρέπει τη σύγκριση δραστικότητας<br />
διαφορετικών ενζύμων).
1) Ειδική δραστικότητα ή ενεργότητα ή καθαρότητα (specific<br />
activity): Οι μονάδες ενζυμικής δραστικότητας ανά mg<br />
πρωτεΐνης, (U/mg πρωτεΐνης) ενζύμου ή ενζυμικού<br />
παρασκευάσματος (γιατί συνήθως χειριζόμαστε ακάθαρτα<br />
παρασκευάσματα, π.χ. ειδική δραστικότητα ενζυμικού<br />
παρασκευάσματος).<br />
• Δηλαδή, η ειδική δραστικότητα καθορίζεται από το ποσό<br />
του προϊόντος που παράγεται ανά μονάδα βάρους ενζύμου.<br />
• Επειδή το ένζυμο που μελετάται συνήθως δεν είναι 100%<br />
καθαρό, αναγκαστικά χρησιμοποιούνται αυθαίρετες μονάδες,<br />
ανάλογα με τις πειραματικές συνθήκες (γιαναπροκύπτουν<br />
εύχρηστες αριθμητικές τιμές).<br />
• Έτσι η καταλυτική δράση του ενζύμου εκφράζεται σε<br />
αυθαίρετες μονάδες ενζυμικής δραστικότητας ανά mg<br />
πρωτεΐνης (ή σπανιότεραανά100.000g, θεωρώντας ότι το<br />
μοριακό βάρος των ενζύμων είναι περίπου 100.000).
2) Μοριακή δραστικότητα (molecular activity): Οι μονάδες<br />
ενζυμικής δραστικότητας ανά μmol ενζύμου, σε βέλτιστες<br />
συνθήκες συγκέντρωσης υποστρώματος.<br />
• Δηλαδή είναι ο αριθμός των μορίων υποστρώματος που<br />
αλλοιώνονται σε 1 min από 1 μόριο ενζύμου.<br />
• Με άλλα λόγια, είναι το ποσό του προϊόντος που<br />
παράγεται από ένα μόριο ενζύμου.<br />
• Από την ενζυμική κινητική είναι γνωστό ότι<br />
Vmax = κ 2 [Εt].<br />
Όπου [Εt] είναι η συγκέντρωση του συνολικού ενζύμου.<br />
• Όταν [Εt] = 1, τότε Vmax= κ 2 και μετριέται σε min-1.<br />
• To κ 2 oνομάζεται αριθμός ανακύκλωσης (turnover number)<br />
και δείχνει πόσα μόρια υποστρώματος αλλοιώνονται στη<br />
μονάδα του χρόνου από ένα μόριο ενζύμου.<br />
• Οι τιμές που έχουν ανακοινωθεί για τους αριθμούς<br />
ανακύκλωσης των διαφόρων ενζύμων κυμαίνονται από 6<br />
έως 17x10 6 .
3) Δραστικότητα καταλυτικού κέντρου: Οτρόποςέκφρασης<br />
της καταλυτικής δράσης ενός ενζύμου, στην περίπτωση που<br />
το ένζυμο έχει<br />
• προσθετική ομάδα ή καταλυτικό κέντρο και<br />
• εφ’ όσον η συγκέντρωση αυτών μπορεί να<br />
προσδιορισθεί,<br />
και ισούται με τον αριθμό των μορίων του υποστρώματος<br />
που αλλοιώνονται ανά 1min, από 1 προσθετική<br />
ομάδα/καταλυτικό κέντρο.
Μέθοδοι ποσοτικού προσδιορισμού της ενζυμικής<br />
δράσης<br />
Οι πιο σημαντικές κατηγορίες μεθόδων που χρησιμοποιούνται<br />
σήμερα για τον προσδιορισμό της καταλυτικής<br />
δράσης των ενζύμων είναι οι εξής:<br />
• Φασματοφωτομετρικές μέθοδοι: Πολλά υποστρώματα και<br />
προϊόντα μιας ενζυμικής αντίδρασης απορροφούν την<br />
ακτινοβολία στην περιοχή του ορατού ήτουυπεριώδους<br />
φάσματος.<br />
• Σε αυτές τις περιπτώσεις μπορεί να επιλεγεί ένα μήκος<br />
κύματος, στο οποίο θα παρακολουθείται ποσοτικά είτε η<br />
σταδιακή εξαφάνιση του υποστρώματος, είτε η σταδιακή<br />
εμφάνιση του προϊόντος.<br />
• Όταν κανένα από τα δύο συστατικά της αντίδρασης δεν<br />
απορροφά την ακτινοβολία, ο ποσοτικός προσδιορισμός<br />
μπορεί να πραγματοποιηθεί με τη σύζευξη της<br />
αντίδρασης που μελετάται, με μία άλλη.
Παραδείγματα φασματοφωτομετρικών μεθόδων:<br />
1. Προσδιορισμός γαλακτικής αφυδρογονάσης:<br />
Γαλακτικό οξύ + NAD +<br />
Γαλακτική αφυδρογονάση<br />
Πυροσταφυλικό οξύ + NADH + H +<br />
•Το ΝΑD + απορροφά στα 260nm<br />
•Το ΝΑDΗ στα340 - 260nm.<br />
•Έτσι μπορούμε να μετρήσουμε είτε<br />
• την ελάττωση της συγκέντρωσης του NAD + , είτε<br />
• την αύξηση της συγκέντρωσης του NADH
2. Προσδιορισμός κινάσης 6-φωσφοφρουκτόζης:<br />
6-φωσφοφρουκτόζη<br />
1,6-διφωσφοφρουκτόζη<br />
κινάση της 6-<br />
φωσφοφρουκτόζης<br />
αλδολάση<br />
1,6-διφωσφοφρουκτόζη 3-φωσφογλυκεριναλδεΰδη +<br />
φωσφοδιυδροξυακετόνη<br />
Αυτά τα προϊόντα δεν απορροφούν στο υπεριώδες, αλλά<br />
3-φωσφογλυκεριναλδεΰδη + NAD + + Pi<br />
1,3-διφωσφογλυκερινικό οξύ + NADH<br />
αφυδρογονάση της 3-<br />
φωσφογλυκεριναλδεΰδης<br />
Έτσι, επιτυγχάνονται έμμεσα οι συνθήκες του προηγούμενου<br />
παραδείγματος.
Φθορισμομετρικές μέθοδοι: Είναι πιο ευαίσθητες από τις<br />
φασματοφωτομετρικές μεθόδους.<br />
•Χρησιμοποιείται κυρίως όταν κάποιο από τα συστατικά<br />
της αντίδρασης φθορίζει π.χ. οι φλαβινο-ενώσεις, οι<br />
οποίες φθορίζουν έντονα στις οξειδωμένες τους μορφές.<br />
• Μπορεί ακόμα να εφαρμοσθεί η μέθοδος αυτή μετά από<br />
μετατροπή κάποιου από τα αντιδρώντα συστατικά σε<br />
φθορίζον παράγωγο.<br />
Μανομετρικές μέθοδοι: Βρίσκουν κυρίως εφαρμογή στις<br />
ενζυμικές αντιδράσεις, όπου ένα από τα συστατικά είναι<br />
αέριο.<br />
•Έχουν υιοθετηθεί για τη μελέτη των οξειδασών<br />
(πρόσληψη Ο 2 ), των αποκαρβοξυλασών (αποβολή CΟ 2 )<br />
κ.λπ.<br />
•Οι μανομετρικές μέθοδοι μπορούν να χρησιμοποιηθούν<br />
και στις περιπτώσεις που μία ενζυμική αντίδραση είναι σε<br />
σύζευξη με μία άλλη και ένα από τα συστατικά της είναι<br />
αέριο.
Μέθοδοι με ηλεκτρόδια: Ηχρήσητουηλεκτροδίου υάλου<br />
δίνει τη δυνατότητα να παρακολουθούνται οι αντιδράσεις<br />
κατά τις οποίες απελευθερώνονται όξινα προϊόντα,<br />
• είτε με μέτρηση της πτώσης της τιμής του pΗ<br />
συναρτήσει του χρόνου<br />
• (μειονεκτήματα είναι ότι<br />
• η αλλαγή στο pΗ κατά τη διάρκεια της αντίδρασης<br />
θα επιφέρει και αλλαγή στη δραστικότητα του<br />
ενζύμου<br />
• ο ρυθμός μείωσης του pΗ εξαρτάται επίσης και<br />
από τη ρυθμιστική δύναμη του ενζυμικού<br />
συστήματος)<br />
• είτε με συνεχή και αυτόματη τιτλοδότηση των όξινων<br />
ομάδων με άλκαλι.
Πολωσιμετρικές μέθοδο εφαρμόζονται σε:<br />
•Ένζυμα που δρουν μόνο στο ένα οπτικό ισομερές ενός<br />
υποστρώματος και σ’ αυτές τις περιπτώσεις συνήθως τα<br />
προϊόντα είναι οπτικώς ανενεργά.<br />
•Ένζυμα που δρουν σε ανενεργό το υπόστρωμα και τα<br />
προϊόντα είναι οπτικώς ενεργά.<br />
•Ένζυμα που το υπόστρωμα και τα προϊόντα είναι<br />
ενεργά, με διαφορετικές τιμές στροφικής ικανότητας.<br />
• Σε όλες τις παραπάνω περιπτώσεις, η παρακολούθηση της<br />
ενζυμικής αντίδρασης γίνεται με μέτρηση της ικανότητας<br />
περιστροφής του πολωμένου φωτός συναρτήσει του χρόνου.<br />
• Κλασικό παράδειγμα αποτελεί η αντίδραση<br />
σακχαράση<br />
σακχαρόζη<br />
φρουκτόζη + γλυκόζη<br />
Ησακχαρόζηέχεια D20 = +66,5, ενώ<br />
το μίγμα γλυκόζης και φρουκτόζης έχει α D20 =-39,8.
Χημικές μέθοδοι: Ο βαθμός προόδου πολλών ενζυμικών<br />
αντιδράσεων παρακολουθείται με τη λήψη δειγμάτων σε<br />
διάφορες χρονικές στιγμές και στη συνέχεια με<br />
προσδιορισμό του υποστρώματος ή του προϊόντος, με<br />
χημικές μεθόδους.<br />
•Για παράδειγμα οι φωσφατάσες, φωσφορυλάσες και<br />
νουκλεοτιδάσες, προσδιορίζονται με μέτρηση του ποσού<br />
του ανόργανου φωσφόρου που απελευθερώνεται ή<br />
καταναλώνεται.<br />
•Tο μειονέκτημα αυτών των μεθόδων είναι ότι απαιτούν<br />
περισσότερο χρόνο.<br />
Χρωματογραφικές μέθοδοι: Στις περιπτώσεις που δεν μπορεί<br />
να εφαρμοστεί καμμία από τις παραπάνω μεθόδους:<br />
• Η ανίχνευση του σχηματισμού του προϊόντος μπορεί να<br />
γίνει χρωματογραφικά (το R f ή/και ο χρόνος έκλουσης είναι<br />
ενδείξεις για την ταυτοποίησή του).<br />
• Στη συνέχεια, στις καθαρές ενώσεις μπορεί να γίνει<br />
ποσοτικός προσδιορισμός.
Η χρήση ραδιενεργών ισοτόπων: Είναι ένα χρήσιμο εργαλείο<br />
για την παρακολούθηση των ενζυμικών αντιδράσεων.<br />
• Για παράδειγμα το ραδιενεργό προϊόν ήτοεναπομένον<br />
υπόστρωμα (μετά το διαχωρισμό τους) προσδιορίζεται<br />
ποσοτικά με μετρητή ραδιενέργειας .
ΕΠΙΔΡΑΣΗ ΤΗΣ ΣΥΓΚΕΝΤΡΩΣΗΣ ΤΟΥ ΥΠΟΣΤΡΩΜΑΤΟΣ<br />
ΣΤΗΝ ΤΑΧΥΤΗΤΑ ΤΗΣ ΑΝΤΙΔΡΑΣΗΣ<br />
• Η πλειοψηφία των ενζυμικών αντιδράσεων περιλαμβάνει<br />
περισσότερα από ένα υποστρώματα.<br />
•Ακόμα και στις υδρολάσες, οι οποίες συχνά θεωρούνται<br />
αντιδράσεις ενός υποστρώματος, το δεύτερο υπόστρωμα<br />
(το Η 2 Ο) υπάρχει σε υψηλές συγκεντρώσεις, και το ένζυμο<br />
να είναι κορεσμένο στο Η 2 Ο σε όλες τις χρονικές στιγμές.<br />
• Αν και η περίπτωση των ενζυμικών αντιδράσεων ενός<br />
υποστρώματος είναι σπάνια στα βιοχημικά συστήματα, παρ’<br />
όλ’ αυτά αντιπροσωπεύει ένα χρήσιμο μοντέλο για την<br />
ανάπτυξη της θεωρίας της ενζυμικής κινητικής και τα<br />
περισσότερα συμπεράσματα που εξάγονται βρίσκουν<br />
εφαρμογή και στα πιο πολύπλοκα ενζυμικά συστήματα.<br />
• Για τους λόγους αυτούς, θα αναπτυχθεί κυρίως η κινητική<br />
των ενζυμικών αντιδράσεων με ένα υπόστρωμα πριν<br />
αναφερθούν οι περιπτώσεις των ενζύμων με πολλά<br />
υποστρώματα.
Η συγκέντρωση του υποστρώματος [S], είναι ένας από τους<br />
σημαντικότερους παράγοντες που επιδρούν στην ταχύτητα<br />
[v], των ενζυμικών αντιδράσεων.<br />
• Σε μια πρώτης τάξης μη εvζυμική αvτίδραση, η μεταβoλή<br />
της ταχύτητας, καθώς αυξάvει η συγκέντρωση του<br />
υποστρώματος, είvαι σταθερή και η καμπύλη v = f(S) είvαι<br />
ευθεία.<br />
• Στις περισσότερες<br />
περιπτώσεις σε μια<br />
ενζυμική αντίδραση,<br />
το διάγραμμα της αρχικής<br />
ταχύτητας (ν) συναρτήσει<br />
της συγκέντρωσης<br />
του υποστρώματος<br />
έχει τη μορφή<br />
καμπύλης υπερβολής.
Τα σημεία που προέρχονται από το πείραμα της μέτρησης<br />
της v=f (S), βρίσκονται επάνω σε μια καμπύλη, που αποτελεί<br />
τμήμα της καμπύλης<br />
που αποδίδεται<br />
μαθηματικά σαν<br />
καμπύλη υπερβολής<br />
(χψ
• Αν ορισθούν οι συντεταγμένες (χ 0 , ψ 0 ) της αρχής (Ο) των<br />
αξόνων (χ',ψ' ή v,[S]) του τμήματος της καμπύλης υπερβολής<br />
που αντιστοιχεί στη<br />
γραφική παράσταση<br />
v=f([S]) του ενζυμικού<br />
συστήματος και<br />
θεωρηθεί ότι χ 0 =Κ Μ<br />
και ψ 0 = -Vmax,<br />
τότε ισχύουν τα<br />
ακόλουθα:<br />
• Τυχόν σημείο<br />
Μ (χ'=[S], ψ'= v)<br />
έχει συντεταγμένες<br />
χ=χ'+χ 0 , ψ=ψ΄+ψ 0<br />
·
• Θέτοντας τις τιμές αυτές στην εξίσωση υπερβολής<br />
χψ=σταθερό, προκύπτει (χ'+χ 0 )(ψ΄+ψ 0 )=χ 0 ψ 0 οπότε<br />
ψ'(χ'+χ 0 )=χ 0 ψ 0 -ψ 0 (χ'+χ 0 )=-ψ 0 χ' και λύνοντας ως προς ψ΄<br />
ψ' = -ψ 0 χ'/χ'+χ 0<br />
Αλλά εξ ορισμού:<br />
• το ψ' = v<br />
• το χ' = S<br />
• το ψ 0 = - Vmax και<br />
• το χ 0 = K M<br />
Και η προηγούμενη<br />
εξίσωση λαμβάνει<br />
τη γνωστή μορφή<br />
της εξίσωσης<br />
Michaelis-Menten,<br />
v = Vmax[S]/[S]+K M
Στις ενζυμικές αντιδράσεις ακολουθείται κινητική κορεσμού.<br />
•Σε χαμηλές [S], η v εξαρτάται τόσο από τη [S] όσο και από<br />
τη [E].<br />
• Από μία τιμή [S] και μετά (όταν το Ε έχει κορεσθεί από το S)<br />
η v είναι ανεξάρτητη<br />
από τη [S] και<br />
εξαρτάται μόνο<br />
από τη [E].<br />
• Δηλαδή, σε υψηλές<br />
τιμές [S], το E έχει<br />
κορεσθεί από<br />
το S και η v έχει<br />
αποκτήσει τη<br />
μέγιστη τιμή της<br />
το Vmax.
• Tο 1902, οι Βrown και Ηenri πρότειναν ότι το ένζυμο<br />
δημιουργεί αρχικά ένα σύμπλοκο με το υπόστρωμα, το<br />
οποίο διασπάται στη συνέχεια σε ένζυμο και στα<br />
προϊόντα της αντίδρασης (Ρ)<br />
Ε + S ES (1) και ES → E + P (2)<br />
όπου οι κ 1 , κ -1 και κ 2 σταθερές ταχύτητας,<br />
με διαστάσεις Μ -1 sec -1 για την κ 1 και<br />
sec -1 για τις κ -1 και κ 2 .<br />
• Tο 1913, οι Μichaelis - Μenten απέδωσαν με μαθηματικό<br />
τρόπο το μηχανισμό δράσης των ενζύμων, στηριζόμενοι<br />
στην ιδέα της δημιουργίας του ενδιάμεσου συμπλόκου<br />
ενζύμου-υποστρώματος.
Θεωρία των Μichaelis - Μenten<br />
(παραδοχή αποκατάστασης ισορροπίας)<br />
• Βασίζεται στη γενική παραδοχή ότι έχει αποκατασταθεί μια<br />
ισορροπία στο σύστημα.<br />
•Oι σημαντικέςπαραδοχές είναιοιεξής:<br />
κ 1<br />
• Η αντίδραση μεταξύ του Ε και του S, Ε + S ES (1)<br />
παραμένει σε ισορροπία και<br />
• οποιαδήποτε επίδραση της αντίδρασης ES → E + P (2)<br />
κ<br />
στην (1), θεωρείται αμελητέα.<br />
2<br />
• Οι συνθήκες αυτές επιτυγχάνονται όταν ο ρυθμός<br />
διάσπασης του συμπλόκου ΕS προς Ε και S είναι πολύ<br />
μεγαλύτερος από το ρυθμό διάσπασής του σε Ε και Ρ<br />
(δηλαδή κ -1 >>κ 2 ).<br />
• Η συγκέντρωση του ελεύθερου S παραμένει σχεδόν<br />
αμετάβλητη κατά την αρχική περίοδο της αντίδρασης,<br />
και ισούται με τη συγκέντρωση του ολικού S,<br />
δηλαδή [S] = [St].<br />
κ -1
• Η σταθερά διάστασης (Κs) του συμπλόκου ΕS ορίζεται ως<br />
[E][S] κ<br />
K = =<br />
-1<br />
( 3)<br />
S<br />
[ES] κ<br />
Οι διαστάσεις της Κs είναι sec-1/M-1 sec-1=M,<br />
δηλαδή η Κs έχει διαστάσεις συγκέντρωσης.<br />
• Στην (3) ισχύει η σχέση [Ε] = [Εt] - [ΕS] (4)<br />
Η ταχύτητα της συνολικής αντίδρασης, με βάση την<br />
αντίδραση ES → E + P (2) δίνεται από τον τύπο<br />
• Με αντικατάσταση της [Ε] από την εξίσωση (4) στην<br />
εξίσωση (3) λαμβάνεται η κάτωθι εξίσωση (6)<br />
+1<br />
dP<br />
v=<br />
= κ 2 [ ΕS] ( 5<br />
dt<br />
([ E ] - [ES]) [S]<br />
K =<br />
t<br />
( 6 )<br />
S<br />
[ES]<br />
)
• Η εξίσωση (6) αν λυθεί ως προς [ΕS] και αντικατασταθεί<br />
στην εξίσωση (5) λαμβάνεται η εξίσωση (7)<br />
v=<br />
κ<br />
2<br />
[ E<br />
t<br />
[S] + K<br />
][S]<br />
S<br />
• Όταν η [S] είναι πολύ μεγάλη σε σύγκριση με την Κ S<br />
(δηλαδή Κ S /S τείνει στο 0), τότε η v ισούται με το γινόμενο<br />
κ 2 [Εt] και έχει ήδη ορισθεί σαν Vmax.<br />
(<br />
7<br />
)<br />
v<br />
=<br />
κ<br />
2<br />
[ E<br />
t<br />
[S]<br />
][S]<br />
=<br />
κ<br />
2<br />
[<br />
Ε<br />
t<br />
] (<br />
8<br />
)<br />
• Άρα η εξίσωση (8) μπορεί να γραφεί<br />
v=<br />
V<br />
max<br />
[S]<br />
(<br />
9<br />
)<br />
[S] + K<br />
S
• Η σταθερά διάστασης του συμπλόκου (Κ S ) ονομάζεται<br />
σταθερά Μichaelis - Μenten (Κ Μ ) και η εξίσωση (9)<br />
v=<br />
V<br />
max<br />
[S]<br />
( 9 )<br />
[S] + K<br />
S<br />
παίρνει την τελική μορφή της εξίσωσης Μichaelis- Μenten<br />
(10)<br />
v=<br />
V<br />
max<br />
[S]<br />
[S] + K<br />
M<br />
( 10<br />
)
Θεωρία των Βriggs – Ηaldane<br />
(παραδοχή της αποκατάστασης σταθεροποιημένης<br />
κατάστασης)<br />
• Μία διαφορετική μαθηματική προσέγγιση για τον τρόπο<br />
δράσης των ενζύμων, αποδόθηκε το 1925 από τους Βriggs<br />
και Ηaldane.<br />
•Η θεωρίατουςβασίστηκε<br />
στην παραδοχή<br />
ότι σε κάθε χρονική<br />
στιγμή, ο ρυθμόςσχηματισμού<br />
και διάσπασης<br />
του συμπλόκου<br />
ΕS είναι σχεδόν ίσος,<br />
έτσι ώστε να αποκαθίσταται<br />
μια δυναμική<br />
ισορροπία<br />
(steady state).
• Με βάση τη θεωρία αποκατάστασης σταθεροποιημένης<br />
κατάστασης<br />
κ 1 κ 2<br />
Ε + S ES → E + P<br />
ισχύει ότι<br />
κ -1<br />
d [ES]<br />
= κ 1[E][S] -κ<br />
-1[ES] -κ<br />
2 [ES] = 0 ( 11)<br />
dt<br />
[Ε] = [Εt] - [ΕS] (4)<br />
dP<br />
= [ S] ( )<br />
v=<br />
κ 2 Ε 5<br />
dt<br />
Λύνοντας την (4) ως προς [Ε] και αντικαθιστώντας την τιμή<br />
αυτή στην (11), η οποία λύνεται στη συνέχεια ως προς [ES]<br />
και αντικαθιστώντας την τιμή αυτή στην (5) προκύπτει η<br />
σχέση
v=<br />
κ 2 [ Et<br />
] κ 1[S]<br />
κ [S] + κ + κ<br />
1<br />
-1<br />
2<br />
v=<br />
V max [S]<br />
-1+<br />
[S] +<br />
κ κ<br />
κ<br />
1<br />
2<br />
( 12 )<br />
• Ονομάζοντας το κλάσμα (κ -1<br />
+ κ 2<br />
)/κ 1<br />
= Κ Μ<br />
, η τελευταία<br />
εξίσωση είναι ίδια με την εξίσωση των Μichaelis-Μenten, με<br />
εξαίρεση ότι η Κ S<br />
έχει αντικατασταθεί από την Κ Μ<br />
όπου<br />
κ + κ κ<br />
K = − 1 2<br />
= K +<br />
2<br />
(13<br />
M S<br />
κ<br />
1<br />
κ<br />
+1<br />
)<br />
• Η Κ Μ έχει διαστάσεις συγκέντρωσης διότι<br />
• τόσο η Κ S<br />
• όσο και ο λόγος κ 2 /κ 1 =sec -1 /M -1 sec -1 =M,<br />
έχουν διαστάσεις συγκέντρωσης.
Φυσική σημασία των σταθερών Κ Μ και Vmax<br />
• Για τον πλήρη ορισμό της φυσικής σημασίας της σταθεράς<br />
Κ Μ πρέπει να είναι γνωστές οι τιμές των σταθερών ταχύτητας<br />
αφού με βάση την εξίσωση<br />
κ + κ κ<br />
K = − 1 2<br />
= K +<br />
2<br />
(13<br />
M S<br />
κ<br />
μπορεί να ισχύουν οι σχέσεις:<br />
• κ -1 >> κ 2<br />
• κ -1 = κ 2<br />
• κ -1 < < κ 2<br />
1<br />
κ<br />
+1<br />
)
Κ Μ =<br />
κ -1 + κ 2<br />
κ 1 κ 2<br />
Ε + S ES → E + P<br />
κ -1<br />
κ +1<br />
Κ S =<br />
κ -1<br />
κ +1<br />
• Αν κ -1 > > κ 2 , τότε η εξίσωση απλοποιείται και<br />
ηΚ Μ ισούται με την Κ S και<br />
ισχύει η παραδοχή της αποκατάστασης ισορροπίας που<br />
έγινε στη θεωρία Μichaelis-Μenten,<br />
δηλαδή είναι η σταθερά διάστασης του συμπλόκου<br />
ενζύμου- υποστρώματος.<br />
• Αν κ -1 = κ 2 , τότε η εξίσωση παραμένει ως έχει και<br />
ηΚ Μ έχει την τιμή που βγαίνει από την παραδοχή της<br />
αποκατάστασης σταθεροποιημένης κατάστασης.
κ 1<br />
Ε + S ES → E + P<br />
κ -1<br />
κ -1 + κ 2<br />
κ -1<br />
Κ Μ =<br />
Κ S =<br />
κ<br />
κ +1<br />
+1<br />
• Αν κ -1 < < κ 2 , τότε η Κ Μ ισούται με κ 2 /κ 1 δηλαδή δεν είναι<br />
παρά ο λόγος δύο σταθερών ταχύτητας των δύο διαδοχικών<br />
και πρακτικά μη αμφίδρομων αντιδράσεων<br />
κ 1 κ 2<br />
E + S ES E + P<br />
όπου το αποτέλεσμα εξαρτάται πλέον από τη σχέση μεταξύ<br />
κ 2 και κ 1 δηλαδή αν<br />
• κ 2 κ 1<br />
κ 2
κ 1 κ 2<br />
E + S ES E + P Κ Μ = κ 2<br />
κ 1<br />
• Αν κ 2
κ 1 κ 2<br />
E + S ES E + P Κ Μ = κ 2<br />
κ 1<br />
• Αν όμως κ 2 >κ 1 τότε δεν μπορεί ν’ αποκατασταθεί ούτε<br />
δυναμική ισορροπία, διότι το σύμπλοκο ES μόλις<br />
σχηματισθεί διασπάται (δηλαδή η ταχύτητα διάσπασης είναι<br />
μεγαλύτερη από την ταχύτητα σχηματισμού του ES).<br />
• Εδώ ισχύουν τότε οι ίδιοι νόμοι που ισχύουν σε αντίστοιχες<br />
μη καταλυτικές χημικές αντιδράσεις που συντελούνται σε δύο<br />
ή περισσότερα στάδια (με το σχηματισμό βραχύβιων<br />
ενδιάμεσων προϊόντων), οπότε η ταχύτητα της συνολικής<br />
αντίδρασης ισούται με την ταχύτητα του βραδύτερου<br />
σταδίου.<br />
• Στην περίπτωσή μας αυτό (ταχύτητα του βραδύτερου<br />
σταδίου) είναι η πρώτη αντίδραση και<br />
v= κ 1 [E][S]
κ 1 κ 2<br />
E + S ES E + P Κ Μ = κ 2<br />
κ 1<br />
• Αφού, όπως αναφέρθηκε, το ES είναι βραχύβιο, πρακτικά<br />
όλοτοένζυμοείναισεκάθεχρονικήστιγμήδιαθέσιμογια<br />
αντίδραση, δηλαδή [E] = [Et] –[ES] = [Et].<br />
• Οπότε η v= κ 1 [E][S] γίνεται v=κ 1 [Et].<br />
• Αφού όμως και το [Et]=σταθερό, θα είναι και το v=σταθερό<br />
(δηλαδή dv/dt =0), οπότε η αντίδραση έχει τη μορφή<br />
αντίδρασης ψευδομηδενικής τάξης μέχρις ότου το [S]<br />
ελαττωθεί σημαντικά.<br />
• Όταν [S]
• Με άλλα λόγια, η εξίσωση Michaelis-Menten μπορεί να<br />
θεωρηθεί μερική περίπτωση της γενικής εξίσωσης της<br />
Θεωρία των Βriggs – Ηaldane που ισχύει όταν η ταχύτητα<br />
διάσπασης του [ES] προς E+P είναι αμελητέα (δηλαδή η κ 2<br />
είναι πολύ μικρότερη της κ -1 ).<br />
• Από τα παραπάνω φαίνεται ότι αν η ταχύτητα μιας<br />
ενζυμικής αντίδρασης ανταποκρίνεται στην εξίσωση<br />
Μichaelis-Μenten, δε σημαίνει ότι ανταποκρίνεται και στο<br />
πρότυπό της, δηλαδή ότι η Κ Μ είναι σταθερά διάστασης του<br />
συμπλόκου και ισούται με την Κ S .<br />
Vmax<br />
• Αν στις εξισώσεις των δύο θεωριών<br />
v=<br />
δώσουμε v=Vmax/2, προκύπτει ότι<br />
[S]+K<br />
Κ Μ =[S], αλλά και το αντίστροφο.<br />
• Δηλαδή η Κ Μ ισούται με εκείνη τη<br />
συγκέντρωση υποστρώματος, στην<br />
οποία η ταχύτητα της αντίδρασης έχει<br />
τη μισή τιμή της μέγιστης ταχύτητας.<br />
[S]<br />
M
• Η σταθερά Κ Μ όπως φαίνεται<br />
• έχει μονάδες συγκέντρωσης και<br />
• αποτελεί χαρακτηριστική σταθερά του ενζύμου για<br />
συγκεκριμένο υπόστρωμα.<br />
• Επιπλέον,<br />
• όσο μεγαλύτερη είναι η τιμή της Κ Μ , δηλαδή<br />
(όσο περισσότερο ποσό υποστρώματος απαιτείται για να<br />
αποκτήσει η<br />
αντίδραση<br />
v = Vmax/2)<br />
• τόσο μικρότερη<br />
είναι<br />
ητάσησύνδεσης<br />
(χημική συγγένεια)<br />
μεταξύ Ε και S και<br />
αντίστροφα.
• Τo Κ Μ είvαι σπoυδαία σταθερά χαρακτηριστική τoυ<br />
εvζύμoυ,<br />
• έστω και αv δεv είvαι γvωστή η ακριβής σημασία της,<br />
• αφoύ καθoρίζει τηv πoσoτική εξάρτηση τoυ S από τo<br />
E.<br />
• Η Κ Μ αποτελεί λοιπόν ποσοτική έκφραση της αντίδρασης<br />
σχηματισμού του συμπλόκου ΕS<br />
Ε + S → ΕS<br />
δείχνει την χημική συγγένεια του Ε και S και είναι ανεξάρτητη<br />
από τη συγκέντρωση του ενζύμου.<br />
• Η Vmax αποτελεί ποσοτική έκφραση της αντίδρασης<br />
διάσπασης του ΕS<br />
ΕS → Ε+ Ρ<br />
και εξαρτάται από τη συγκέντρωση του ενζύμου.
• Οι παράγοντες που επηρεάζουν την αρχική ταχύτητα των<br />
ενζυμικών αντιδράσεων, επιδρώντας είτε<br />
• στο σχηματισμό του συμπλόκου, είτε<br />
• στη διάσπασή του, είτε<br />
• και στα δύο,<br />
μεταβάλλουν τις τιμές των Κ Μ και Vmax.<br />
• Όταν είναι γνωστή η μοριακή συγκέντρωση του ενζύμου,<br />
τότε το Vmax εκφράζεται σε μονάδες ποσότητας (mol/L=M)<br />
αντιδρώντος υποστρώματος (ή σχηματιζόμενου προϊόντος)<br />
ανά μονάδα χρόνου, για συγκεκριμένη συγκέντρωση<br />
ενζύμου, δηλαδή έχει μονάδες Μ.min -1 .<br />
• Αν στην εξίσωση Vmax = κ 2 [Εt] θεωρηθεί ότι η [Εt]=1mol/L,<br />
προκύπτει ότι Vmax =κ 2 (min -1 ).<br />
• Για το λόγο αυτό, το κ 2 ονομάζεται αριθμός ανακύκλωσης<br />
και δηλώνει πόσα μόρια υποστρώματος αλλοιώνονται από<br />
ένα μόριο ενζύμου στη μονάδα του χρόνου.<br />
• Ο αριθμός ανακύκλωσης αποτελεί επίσης χαρακτηριστική<br />
σταθερά του ενζύμου για συγκεκριμένο υπόστρωμα
Συνόψιση των παραδοχών για την παραγωγή<br />
της εξίσωσης Μichaelis-Μenten<br />
1. Η δημιουργία του ενδιάμεσου συμπλόκου ΕS. Η υπόθεση<br />
αυτή έχει μεγάλη σημασία στηv κιvητική τωv εvζυμικώv<br />
αvτιδράσεωv και στηv καταvόηση τoυ μηχαvισμoύ<br />
αvαστoλής.<br />
2. Το ένζυμο αντιδρά με ένα μόνο υπόστρωμα, δηλαδή το S<br />
συνδέεται σε ένα μόνο σημείο με το Ε. Αν το ένζυμο αντιδρά<br />
με n μόρια υποστρώματος, γίνεται η αντίδραση<br />
nS + E → ESn → E + nP<br />
και η ταχύτητα αντίδρασης ισούται με<br />
[S]<br />
max<br />
n<br />
[S]<br />
• Η ταχύτητα της αvτίδρασης θα παίρvει<br />
τηv τιμή Vmax/2 όταv ησυγκέvτρωση τoυ υπoστρώματoς<br />
γίvει [S]=K M<br />
1/n<br />
.<br />
• Η γραφική παράσταση 1/v=f(1/S) δεv είvαι ευθεία,<br />
ν<br />
=<br />
V<br />
+<br />
K<br />
n<br />
M
3. Στην ημιαντίδραση Ε+S ΕS θεωρείται ότι<br />
αποκαθίσταται ισορροπία, ενώ στη δεύτερη ημιαντίδραση<br />
ES→E+P, ότι το ΕS διασπάται γρήγορα προς ένζυμο και<br />
προϊόντα.<br />
• Δηλαδή γίvεται η παραδoχή ότι η ταχύτητα σχηματισμoύ<br />
τoυ ES και η ταχύτητα διάσπασης τoυ ES πρoς ΕκαιS είvαι<br />
πoλύ μεγαλύτερη από τηv ταχύτητα διάσπασης τoυ ES πρoς<br />
ΕκαιΡ.<br />
• Όλα τα αvωτέρω συvoψίζovται στηv υπόθεση ότι τo Κ Μ<br />
είvαι η σταθερά διάστασης τoυ ES, πράγμα όχι πάvτα oρθό,<br />
όπως φαίvεται από τη διερεύvηση της σχέσης (13).<br />
• Διότι οι Brigg-Haldane δεν διατήρησαν αυτήν την<br />
παραδοχή, αλλά δέχθηκαν την σταθεροποιημένη κατάσταση<br />
ισοζυγίου, που συμπεριλαμβάνει και την ημιαντίδραση<br />
διάσπασηςπροςΡ.<br />
κ 1 κ 2<br />
E + S ES → Ρ<br />
κ -1
4. Ενώ η συγκέντρωση του συνολικού ενζύμου ορίζεται σαν<br />
[Εt] = [Ε] + [ΕS]<br />
η συγκέντρωση του ελεύθερου υποστρώματος θεωρείται ίση<br />
με τη συγκέντρωση του ολικού υποστρώματος<br />
[St]=[S]<br />
• Η παραδοχή αυτή μπορεί να γίνει, γιατί ησχετική<br />
συγκέντρωση του υποστρώματος ως προς το ένζυμο είναι<br />
πολύ μεγάλη και η δεσμευμένη ποσότητα του υποστρώματος<br />
στο ένζυμο, μπορεί να θεωρηθεί σαν αμελητέα.<br />
• Αν δεν γίνει αυτή η παραδοχή, τότε στον υπολογισμό της<br />
συγκέντρωσης του ΕS θα περιλαμβάνεται εκθετική εξίσωση.
5. Στην περιγραφή των σταδίων που διακρίνονται σε μια<br />
ενζυμική αντίδραση, δεν περιλαμβάνεται η αντίδραση<br />
κ κ 1 2 κ 3<br />
Ε + S ES EP →<br />
κ κ -1 -2<br />
E + P<br />
δηλαδή έγινε η παραδοχή ότι το κ 3 είναι πολύ μεγαλύτερο<br />
από το κ -2 και το κ 2 .<br />
• Αυτό συμβαίvει πoλλές φoρές, χωρίς vα είvαι γvωστό ότι<br />
γίvεται στηv εvζυμική αvτίδραση.<br />
• Αv δεv ισχύει αυτή η υπόθεση, τότε τα Vmax και Κ Μ δίvovται<br />
από τoυς ακόλoυθoυς πoλύπλoκoυς τύπoυς και είvαι πoλύ<br />
δύσκoλo vα εξηγηθεί η φυσική έvvoια τoυ Κ Μ .<br />
V<br />
max<br />
=<br />
κ<br />
κ<br />
2<br />
3<br />
κ<br />
+<br />
2<br />
κ<br />
[E ]<br />
-2<br />
t<br />
+<br />
κ<br />
3<br />
K<br />
M<br />
=<br />
κ<br />
-1<br />
κ<br />
κ<br />
1<br />
-2<br />
(<br />
κ<br />
+<br />
2<br />
κ<br />
+<br />
-1<br />
κ<br />
κ<br />
-2<br />
3<br />
+<br />
+<br />
κ<br />
κ<br />
3<br />
2<br />
κ<br />
)<br />
3
6. Tέλος, δε λαμβάνεται υπόψη η αντιστρεπτότητα της<br />
αντίδρασης, αν και είναι γνωστό ότι τα ένζυμα καταλύουν<br />
συνήθως την αντίδραση και προς τις δύο κατευθύνσεις.<br />
• Γι’ αυτό το λόγο, έχει ήδη αναφερθεί ότι η ταχύτητα που<br />
προσδιορίζεται είναι η αρχική, δηλαδή ότι η συγκέντρωση<br />
των προϊόντων θεωρείται αμελητέα.<br />
• Σε αντίθετη περίπτωση, η ταχύτητα της αvτίδρασης θα είvαι<br />
συvάρτηση τωv<br />
•Vmax, K M , [S], αλλά και των<br />
• [P], V'max και Κ' Μ (πρoς τηv αvτίθετη κατεύθυvση).