

Bat Echolocation Researc h - Bat Conservation International
Bat Echolocation Researc h - Bat Conservation International
Bat Echolocation Researc h - Bat Conservation International

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
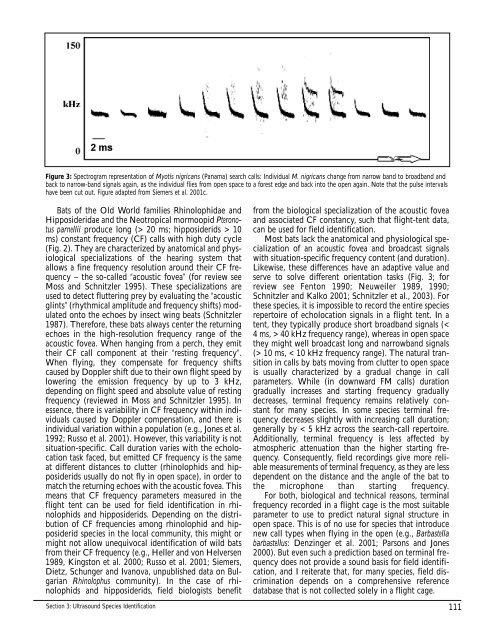
Figure 3: Spectrogram representation of Myotis nigricans (Panama) search calls: Individual M. nigricans change from narrow band to broadband and<br />
back to narrow-band signals again, as the individual flies from open space to a forest edge and back into the open again. Note that the pulse intervals<br />
have been cut out. Figure adapted from Siemers et al. 2001c.<br />
<strong>Bat</strong>s of the Old World families Rhinolophidae and<br />
Hipposideridae and the Neotropical mormoopid Pteronotus<br />
parnellii produce long (> 20 ms; hipposiderids > 10<br />
ms) constant frequency (CF) calls with high duty cycle<br />
(Fig. 2). They are characterized by anatomical and physiological<br />
specializations of the hearing system that<br />
allows a fine frequency resolution around their CF frequency<br />
– the so-called “acoustic fovea” (for review see<br />
Moss and Schnitzler 1995). These specializations are<br />
used to detect fluttering prey by evaluating the “acoustic<br />
glints” (rhythmical amplitude and frequency shifts) modulated<br />
onto the echoes by insect wing beats (Schnitzler<br />
1987). Therefore, these bats always center the returning<br />
echoes in the high-resolution frequency range of the<br />
acoustic fovea. When hanging from a perch, they emit<br />
their CF call component at their “resting frequency”.<br />
When flying, they compensate for frequency shifts<br />
caused by Doppler shift due to their own flight speed by<br />
lowering the emission frequency by up to 3 kHz,<br />
depending on flight speed and absolute value of resting<br />
frequency (reviewed in Moss and Schnitzler 1995). In<br />
essence, there is variability in CF frequency within individuals<br />
caused by Doppler compensation, and there is<br />
individual variation within a population (e.g., Jones et al.<br />
1992; Russo et al. 2001). However, this variability is not<br />
situation-specific. Call duration varies with the echolocation<br />
task faced, but emitted CF frequency is the same<br />
at different distances to clutter (rhinolophids and hipposiderids<br />
usually do not fly in open space), in order to<br />
match the returning echoes with the acoustic fovea. This<br />
means that CF frequency parameters measured in the<br />
flight tent can be used for field identification in rhinolophids<br />
and hipposiderids. Depending on the distribution<br />
of CF frequencies among rhinolophid and hipposiderid<br />
species in the local community, this might or<br />
might not allow unequivocal identification of wild bats<br />
from their CF frequency (e.g., Heller and von Helversen<br />
1989, Kingston et al. 2000; Russo et al. 2001; Siemers,<br />
Dietz, Schunger and Ivanova, unpublished data on Bulgarian<br />
Rhinolophus community). In the case of rhinolophids<br />
and hipposiderids, field biologists benefit<br />
Section 3: Ultrasound Species Identification<br />
from the biological specialization of the acoustic fovea<br />
and associated CF constancy, such that flight-tent data,<br />
can be used for field identification.<br />
Most bats lack the anatomical and physiological specialization<br />
of an acoustic fovea and broadcast signals<br />
with situation-specific frequency content (and duration).<br />
Likewise, these differences have an adaptive value and<br />
serve to solve different orientation tasks (Fig. 3; for<br />
review see Fenton 1990; Neuweiler 1989, 1990;<br />
Schnitzler and Kalko 2001; Schnitzler et al., 2003). For<br />
these species, it is impossible to record the entire species<br />
repertoire of echolocation signals in a flight tent. In a<br />
tent, they typically produce short broadband signals (<<br />
4 ms, > 40 kHz frequency range), whereas in open space<br />
they might well broadcast long and narrowband signals<br />
(> 10 ms, < 10 kHz frequency range). The natural transition<br />
in calls by bats moving from clutter to open space<br />
is usually characterized by a gradual change in call<br />
parameters. While (in downward FM calls) duration<br />
gradually increases and starting frequency gradually<br />
decreases, terminal frequency remains relatively constant<br />
for many species. In some species terminal frequency<br />
decreases slightly with increasing call duration;<br />
generally by < 5 kHz across the search-call repertoire.<br />
Additionally, terminal frequency is less affected by<br />
atmospheric attenuation than the higher starting frequency.<br />
Consequently, field recordings give more reliable<br />
measurements of terminal frequency, as they are less<br />
dependent on the distance and the angle of the bat to<br />
the microphone than starting frequency.<br />
For both, biological and technical reasons, terminal<br />
frequency recorded in a flight cage is the most suitable<br />
parameter to use to predict natural signal structure in<br />
open space. This is of no use for species that introduce<br />
new call types when flying in the open (e.g., Barbastella<br />
barbastellus: Denzinger et al. 2001; Parsons and Jones<br />
2000). But even such a prediction based on terminal frequency<br />
does not provide a sound basis for field identification,<br />
and I reiterate that, for many species, field discrimination<br />
depends on a comprehensive reference<br />
database that is not collected solely in a flight cage.<br />
111













