Dynamics of glutamatergic signaling in the mushroom ... - HAL - ESPCI
Dynamics of glutamatergic signaling in the mushroom ... - HAL - ESPCI
Dynamics of glutamatergic signaling in the mushroom ... - HAL - ESPCI

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
S<strong>in</strong>akevitch et al. Neural Development 2010, 5:10<br />
http://www.neuraldevelopment.com/content/5/1/10<br />
Page 13 <strong>of</strong> 20<br />
be seen <strong>in</strong> <strong>the</strong> calyx and pedunculus presented <strong>in</strong> Figure<br />
7C. GFP immunoreactivity associated with dendritic<br />
fields <strong>in</strong> <strong>the</strong> calyx and axons <strong>in</strong> <strong>the</strong> lobes is dramatically<br />
reduced. In addition, any bright axonal branches and<br />
processes have vanished, compared to <strong>the</strong> sta<strong>in</strong><strong>in</strong>g<br />
observed <strong>in</strong> newly eclosed flies. Such a dramatic change<br />
suggests that DmGluRA expression <strong>in</strong> a/bc neuronsis<br />
transient and does not persist for long after eclosion.<br />
The immunoreactivity still detectable at 10 days might<br />
be <strong>the</strong> remnant <strong>of</strong> earlier expression and accumulation<br />
<strong>of</strong> <strong>the</strong> stable GFP prote<strong>in</strong> when <strong>the</strong> DmGluRA promoter<br />
was still active.<br />
High ubiquit<strong>in</strong> levels <strong>in</strong> last-born Kenyon cells <strong>of</strong> young<br />
eclosed flies<br />
Ubiquit<strong>in</strong> is a small prote<strong>in</strong> that b<strong>in</strong>ds covalently to specific<br />
prote<strong>in</strong>s and ei<strong>the</strong>r marks <strong>the</strong>m for degradation by<br />
<strong>the</strong> proteasome or modifies <strong>the</strong>ir activity [72]. Substrate<br />
prote<strong>in</strong> ubiquit<strong>in</strong>ation plays important roles <strong>in</strong> neuronal<br />
differentiation [73,74] and synaptic plasticity [75,76]. We<br />
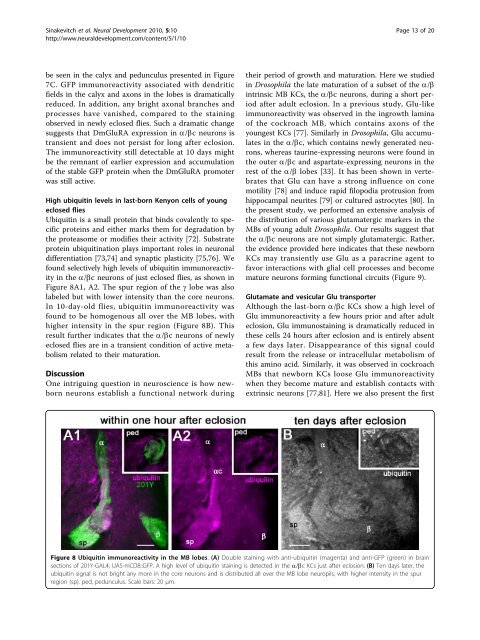
found selectively high levels <strong>of</strong> ubiquit<strong>in</strong> immunoreactivity<br />
<strong>in</strong> <strong>the</strong> a/bc neurons <strong>of</strong> just eclosed flies, as shown <strong>in</strong><br />
Figure 8A1, A2. The spur region <strong>of</strong> <strong>the</strong> g lobe was also<br />
labeled but with lower <strong>in</strong>tensity than <strong>the</strong> core neurons.<br />
In 10-day-old flies, ubiquit<strong>in</strong> immunoreactivity was<br />
found to be homogenous all over <strong>the</strong> MB lobes, with<br />
higher <strong>in</strong>tensity <strong>in</strong> <strong>the</strong> spur region (Figure 8B). This<br />
result fur<strong>the</strong>r <strong>in</strong>dicates that <strong>the</strong> a/bc neurons<strong>of</strong>newly<br />
eclosed flies are <strong>in</strong> a transient condition <strong>of</strong> active metabolism<br />
related to <strong>the</strong>ir maturation.<br />
Discussion<br />
One <strong>in</strong>trigu<strong>in</strong>g question <strong>in</strong> neuroscience is how newborn<br />
neurons establish a functional network dur<strong>in</strong>g<br />
<strong>the</strong>ir period <strong>of</strong> growth and maturation. Here we studied<br />
<strong>in</strong> Drosophila <strong>the</strong> late maturation <strong>of</strong> a subset <strong>of</strong> <strong>the</strong> a/b<br />
<strong>in</strong>tr<strong>in</strong>sic MB KCs, <strong>the</strong> a/bc neurons, dur<strong>in</strong>g a short period<br />
after adult eclosion. In a previous study, Glu-like<br />
immunoreactivity was observed <strong>in</strong> <strong>the</strong> <strong>in</strong>growth lam<strong>in</strong>a<br />
<strong>of</strong> <strong>the</strong> cockroach MB, which conta<strong>in</strong>s axons <strong>of</strong> <strong>the</strong><br />
youngest KCs [77]. Similarly <strong>in</strong> Drosophila, Glu accumulates<br />
<strong>in</strong> <strong>the</strong> a/bc, which conta<strong>in</strong>s newly generated neurons,<br />
whereas taur<strong>in</strong>e-express<strong>in</strong>g neurons were found <strong>in</strong><br />
<strong>the</strong> outer a/bc and aspartate-express<strong>in</strong>g neurons <strong>in</strong> <strong>the</strong><br />
rest <strong>of</strong> <strong>the</strong> a/b lobes [33]. It has been shown <strong>in</strong> vertebrates<br />
that Glu can have a strong <strong>in</strong>fluence on cone<br />
motility [78] and <strong>in</strong>duce rapid filopodia protrusion from<br />
hippocampal neurites [79] or cultured astrocytes [80]. In<br />
<strong>the</strong> present study, we performed an extensive analysis <strong>of</strong><br />
<strong>the</strong> distribution <strong>of</strong> various <strong>glutamatergic</strong> markers <strong>in</strong> <strong>the</strong><br />
MBs <strong>of</strong> young adult Drosophila. Our results suggest that<br />
<strong>the</strong> a/bc neurons are not simply <strong>glutamatergic</strong>. Ra<strong>the</strong>r,<br />
<strong>the</strong> evidence provided here <strong>in</strong>dicates that <strong>the</strong>se newborn<br />
KCs may transiently use Glu asaparacr<strong>in</strong>eagentto<br />
favor <strong>in</strong>teractions with glial cell processes and become<br />
mature neurons form<strong>in</strong>g functional circuits (Figure 9).<br />
Glutamate and vesicular Glu transporter<br />
Although <strong>the</strong> last-born a/bc KCs show a high level <strong>of</strong><br />
Glu immunoreactivity a few hours prior and after adult<br />
eclosion, Glu immunosta<strong>in</strong><strong>in</strong>g is dramatically reduced <strong>in</strong><br />
<strong>the</strong>se cells 24 hours after eclosion and is entirely absent<br />
a few days later. Disappearance <strong>of</strong> this signal could<br />
result from <strong>the</strong> release or <strong>in</strong>tracellular metabolism <strong>of</strong><br />
this am<strong>in</strong>o acid. Similarly, it was observed <strong>in</strong> cockroach<br />
MBs that newborn KCs loose Glu immunoreactivity<br />
when <strong>the</strong>y become mature and establish contacts with<br />
extr<strong>in</strong>sic neurons [77,81]. Here we also present <strong>the</strong> first<br />
Figure 8 Ubiquit<strong>in</strong> immunoreactivity <strong>in</strong> <strong>the</strong> MB lobes. (A) Double sta<strong>in</strong><strong>in</strong>g with anti-ubiquit<strong>in</strong> (magenta) and anti-GFP (green) <strong>in</strong> bra<strong>in</strong><br />
sections <strong>of</strong> 201Y-GAL4; UAS-mCD8::GFP. A high level <strong>of</strong> ubiquit<strong>in</strong> sta<strong>in</strong><strong>in</strong>g is detected <strong>in</strong> <strong>the</strong> a/bc KCs just after eclosion. (B) Ten days later, <strong>the</strong><br />
ubiquit<strong>in</strong> signal is not bright any more <strong>in</strong> <strong>the</strong> core neurons and is distributed all over <strong>the</strong> MB lobe neuropils, with higher <strong>in</strong>tensity <strong>in</strong> <strong>the</strong> spur<br />
region (sp). ped, pedunculus. Scale bars: 20 μm.