10. Automatisme et contrôle nerveux de la motricité digestive ...
10. Automatisme et contrôle nerveux de la motricité digestive ...
10. Automatisme et contrôle nerveux de la motricité digestive ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
<strong>10.</strong> <strong>Automatisme</strong> <strong>et</strong> <strong>contrôle</strong> <strong>nerveux</strong> <strong>de</strong> <strong>la</strong> <strong>motricité</strong><br />
<strong>digestive</strong> (intestinale)<br />
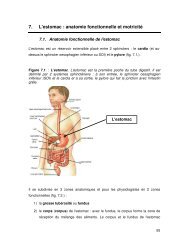
Introduction<br />
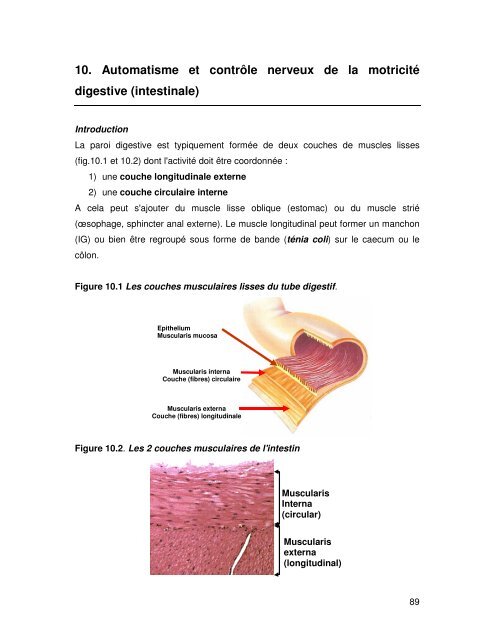
La paroi <strong>digestive</strong> est typiquement formée <strong>de</strong> <strong>de</strong>ux couches <strong>de</strong> muscles lisses<br />
(fig.<strong>10.</strong>1 <strong>et</strong> <strong>10.</strong>2) dont l'activité doit être coordonnée :<br />
1) une couche longitudinale externe<br />
2) une couche circu<strong>la</strong>ire interne<br />
A ce<strong>la</strong> peut s'ajouter du muscle lisse oblique (estomac) ou du muscle strié<br />
(œsophage, sphincter anal externe). Le muscle longitudinal peut former un manchon<br />
(IG) ou bien être regroupé sous forme <strong>de</strong> ban<strong>de</strong> (ténia coli) sur le caecum ou le<br />
côlon.<br />
Figure <strong>10.</strong>1 Les couches muscu<strong>la</strong>ires lisses du tube digestif.<br />
Epithelium<br />
Muscu<strong>la</strong>ris mucosa<br />
Muscu<strong>la</strong>ris interna<br />
Couche (fibres) circu<strong>la</strong>ire<br />
Muscu<strong>la</strong>ris externa<br />
Couche (fibres) longitudinale<br />
Figure <strong>10.</strong>2. Les 2 couches muscu<strong>la</strong>ires <strong>de</strong> l'intestin<br />
Muscu<strong>la</strong>ris<br />
Interna<br />
(circu<strong>la</strong>r)<br />
Muscu<strong>la</strong>ris<br />
externa<br />
(longitudinal)<br />
89
La différence majeure entre un muscle strié <strong>et</strong> un muscle lisse est que ce <strong>de</strong>rnier est<br />
doué d'automatismes alors que l'activité motrice d'un muscle strié est subordonnée<br />
à une comman<strong>de</strong> par le système <strong>nerveux</strong> extrinsèque. Un segment d'intestin isolé <strong>et</strong><br />
p<strong>la</strong>cé dans un liqui<strong>de</strong> <strong>de</strong> survie se contracte régulièrement (fig. <strong>10.</strong>3) ; pour le tube<br />
digestif, le système <strong>nerveux</strong> ne joue qu'un rôle <strong>de</strong> coordination <strong>et</strong> <strong>de</strong> modu<strong>la</strong>tion<br />
<strong>de</strong> l'activité motrice grâce à l'existence <strong>de</strong> plexus <strong>nerveux</strong>.<br />
L'automatisme du tube digestif repose sur l'existence <strong>de</strong> cellules spécialisées : les<br />
cellules interstitielles <strong>de</strong> Cajal (CIC).<br />
Figure <strong>10.</strong>3.: <strong>Automatisme</strong> <strong>de</strong> l'intestin : mise en évi<strong>de</strong>nce in vitro. Un fragment<br />
d'intestin p<strong>la</strong>cé dans un liqui<strong>de</strong> <strong>de</strong> survie se contracte spontanément en <strong>de</strong>hors <strong>de</strong><br />
toute innervation extrinsèque.<br />
<strong>10.</strong>1. Les cellules interstitielles <strong>de</strong> Cajal (CIC)<br />
Les cellules interstitielles <strong>de</strong> Cajal (CIC) sont <strong>de</strong>s cellules différentes <strong>de</strong>s fibres<br />
lisses (elles sont pauvres en éléments contractiles). Elles sont principalement<br />
localisées entre les couches muscu<strong>la</strong>ires longitudinales <strong>et</strong> circu<strong>la</strong>ires au<br />
voisinage <strong>de</strong>s plexus myentériques (fig. <strong>10.</strong>4). Elles développent <strong>de</strong> nombreuses<br />
ramifications interconnectées pour former un réseau. Elles sont connectées aux<br />
cellules lisses par <strong>de</strong>s jonctions serrées ce qui donne à l'ensemble <strong>de</strong>s propriétés<br />
<strong>de</strong> "câble".<br />
90
Figure <strong>10.</strong>4. Les cellules interstitielles <strong>de</strong> Cajal sont à l'origine <strong>de</strong> l'automatisme<br />
<strong>de</strong>s fibres lisses gastro-intestinales. Elles sont situées entre les couches muscu<strong>la</strong>ires<br />
lisses longitudinales <strong>et</strong> circu<strong>la</strong>ires <strong>et</strong> autour <strong>de</strong>s ganglions <strong>de</strong>s plexus myentériques.<br />
Les cellules <strong>de</strong> Cajal ont 4 fonctions majeures :<br />
1) elles sont le siège <strong>de</strong> l'automatisme du tube digestif (pacemaker)<br />
2) elles assurent <strong>la</strong> dépo<strong>la</strong>risation <strong>de</strong>s cellules muscu<strong>la</strong>ires lisses en ouvrant<br />
leurs canaux calciques "voltage dépendant". Ce<strong>la</strong> est à l'origine <strong>de</strong>s on<strong>de</strong>s<br />
lentes (OL) c'est-à-dire <strong>de</strong>s oscil<strong>la</strong>tions régulières du potentiel <strong>de</strong> repos <strong>de</strong>s<br />
fibres lisses. Ces OL déterminent un rythme électrique <strong>de</strong> base (REB) dont<br />
<strong>la</strong> fréquence est propre aux différents segments du tube digestif.<br />
3) elles assurent <strong>la</strong> conduction électronique <strong>de</strong>s OL (propriétés <strong>de</strong> câble).<br />
4) Les CIC situées au cœur même <strong>de</strong>s fibres lisses assurent <strong>la</strong> liaison entre les<br />
motoneurones (excitateurs ou inhibiteurs) <strong>et</strong> les fibres lisses. Les<br />
motoneurones innervent "en passage" <strong>de</strong>s varicosités terminales au contact<br />
avec les CIC.<br />
Les CIC peuvent se regrouper en certaines zones <strong>et</strong> former <strong>de</strong> véritables<br />
pacemaker locaux (comme au niveau <strong>de</strong> <strong>la</strong> gran<strong>de</strong> courbure <strong>de</strong> l'estomac –<br />
91
corpus) (voir fig.7.8). En revanche, tous les segments <strong>de</strong> l'IG possè<strong>de</strong>nt <strong>de</strong>s CIC<br />
qui se répartissent sur l'ensemble <strong>de</strong> sa longueur.<br />
Les CIC sont absentes <strong>de</strong> certaines zones (ex. grosse tubérosité <strong>de</strong> l'estomac)<br />
qui <strong>de</strong> ce fait ne possè<strong>de</strong>nt pas d'activité automatique. De même certaines<br />
pathologies comme l'acha<strong>la</strong>sie seraient dues à <strong>de</strong>s déficits en CIC.<br />
<strong>10.</strong>2. Les on<strong>de</strong>s lentes, le rythme électrique <strong>de</strong> base <strong>et</strong> les<br />
potentiels d'action <strong>de</strong> <strong>la</strong> cellule lisse<br />
L'enregistrement avec <strong>de</strong>s microélectro<strong>de</strong>s intracellu<strong>la</strong>ires <strong>de</strong> l'activité électrique<br />
d'une fibre lisse perm<strong>et</strong> <strong>de</strong> mesurer un potentiel <strong>de</strong> repos <strong>de</strong> l'ordre <strong>de</strong> -50 mV<br />
c'est-à-dire <strong>de</strong> n<strong>et</strong>tement inférieur à celui d'un fibre muscu<strong>la</strong>ire striée ou<br />
cardiaque (-80 mV ).<br />
Sous l'influence <strong>de</strong>s CIC <strong>et</strong> avec une fréquence qui dépend du segment du tube<br />
digestif (3 à 6 par minute pour l'estomac, 12 à 17 par min pour l'IG <strong>et</strong> 9 à 16 sur le<br />
gros intestin chez l'homme), les fibres lisses se dépo<strong>la</strong>risent régulièrement. Les<br />
dépo<strong>la</strong>risation partielles (<strong>de</strong> -50 mV à -40 mV) forment <strong>de</strong>s oscil<strong>la</strong>tions du<br />
potentiel <strong>de</strong> membrane nommées on<strong>de</strong>s lentes <strong>et</strong> leur régu<strong>la</strong>rité donne<br />
naissance à un rythme électrique <strong>de</strong> base (REB) (fig. <strong>10.</strong>5).<br />
La dépo<strong>la</strong>risation partielle va persister sous <strong>la</strong> forme d'un p<strong>la</strong>teau qui peut durer<br />
10 sec au niveau <strong>de</strong> l'estomac. L'on<strong>de</strong> lente n'est pas à l'origine d'une activité<br />
mécanique ; une activité mécanique ne sera vue que si le somm<strong>et</strong> <strong>de</strong> l'on<strong>de</strong> lente<br />
est surchargé <strong>de</strong> potentiels <strong>de</strong> pointe (spikes) encore appelés potentiels<br />
d'action. Les potentiels <strong>de</strong> pointe correspon<strong>de</strong>nt à <strong>de</strong>s dépo<strong>la</strong>risations presque<br />
totales <strong>de</strong> <strong>la</strong> fibre lisse (fig.<strong>10.</strong>5) ; il n'y a pas d'inversion <strong>de</strong> po<strong>la</strong>rité.<br />
La puissance <strong>de</strong> l'activité mécanique est proportionnelle à <strong>la</strong> fréquence <strong>de</strong>s<br />
potentiels <strong>de</strong> pointe. La dépo<strong>la</strong>risation <strong>de</strong> <strong>la</strong> cellule lisse est due à une entrée<br />
<strong>de</strong> calcium <strong>et</strong> non <strong>de</strong> sodium comme pour un axone ou une fibre striée (<strong>la</strong><br />
t<strong>et</strong>rodoxine, un inhibiteur spécifique du canal sodique est sans eff<strong>et</strong> sur <strong>la</strong><br />
dépo<strong>la</strong>risation <strong>de</strong> <strong>la</strong> fibre lisse <strong>digestive</strong>).<br />
92
Figure <strong>10.</strong>5 : <strong>Automatisme</strong> <strong>de</strong> <strong>la</strong> fibre lisse du tube digestif. Le potentiel <strong>de</strong><br />
repos est <strong>de</strong> -50 mV. L'activité automatique <strong>de</strong> <strong>la</strong> fibre se traduit par l'occurrence<br />
<strong>de</strong> dépo<strong>la</strong>risation partielle <strong>de</strong>s cellules intestinales qui forment une succession<br />
d'on<strong>de</strong>s lentes (slow waves), le rythme électrique <strong>de</strong> base (REB). Les<br />
dépo<strong>la</strong>risations ont pour origine les cellules <strong>de</strong> Cajal. Lorsque les potentiels sont<br />
surchargés <strong>de</strong> pointes (spikes), une activité mécanique est déclenchée.<br />
<strong>10.</strong>3. Distribution spatio-temporelle <strong>de</strong>s on<strong>de</strong>s lentes <strong>et</strong><br />
synchronisation <strong>de</strong> l'activité<br />
Sur une section donnée <strong>de</strong> l'IG, on constate une synchronisation du REB entre<br />
les fibres lisses ce qui est à l'origine du caractère annu<strong>la</strong>ire <strong>de</strong> <strong>la</strong> contraction<br />
intestinale (fig.<strong>10.</strong>6). C<strong>et</strong>te synchronisation est due à <strong>la</strong> connectivité assurée par<br />
les CIC. En revanche au niveau du côlon, il n'y a pas <strong>de</strong> synchronisation au<br />
niveau d'une même section ce qui donne <strong>de</strong>s contractions "vermicu<strong>la</strong>ire" plutôt<br />
qu'annu<strong>la</strong>ire (fig.<strong>10.</strong>7). Ces contractions sont plus adaptées au brassage alors<br />
que les contractions annu<strong>la</strong>ires assurent <strong>la</strong> propagation du chyme avec les<br />
contractions péristaltiques.<br />
Figure <strong>10.</strong>6. Propagation synchrone <strong>de</strong>s on<strong>de</strong>s lentes (OL) sur une section<br />
<strong>de</strong> l'intestin grêle. La contraction <strong>de</strong> l'intestin grêle se présente comme un<br />
anneau se dép<strong>la</strong>çant <strong>de</strong> façon orale-aborale.<br />
Temps zéro<br />
5 secon<strong>de</strong>s plus tard<br />
93
Figure <strong>10.</strong>7. Propagation électrotonique <strong>de</strong>s OL : intestin grêle vs. côlon.<br />
Sur une section donnée l'intestin, <strong>la</strong> propagation <strong>de</strong>s OL est synchrone, ce qui<br />
donne une contraction annu<strong>la</strong>ire péristaltique alors que sur le côlon, on observe<br />
une asynchronie dans <strong>la</strong> propagation sur une section donnée ce qui explique <strong>de</strong>s<br />
mouvements <strong>de</strong> type vermicu<strong>la</strong>ires <strong>et</strong> non annu<strong>la</strong>ires.<br />
Intestin grêle Colon<br />
Propagation synchrone<br />
<strong>de</strong>s OL sur une section :<br />
péristaltisme<br />
Sur l'intestin grêle, on constate que <strong>la</strong> fréquence <strong>de</strong>s OL décroît dans le sens<br />
oral-aboral <strong>de</strong> façon discontinue (en marches d'escalier). Ce<strong>la</strong> est dû au fait<br />
que les fréquences intrinsèques <strong>de</strong>s OL en amont sont supérieures à celles <strong>de</strong>s<br />
zones avales. Les zones amont imposent leur rythme à l'aval. Ce n'est que<br />
lorsque <strong>la</strong> différence <strong>de</strong> fréquence intrinsèque entre 2 zones <strong>de</strong>vient trop<br />
importante qu'il y aura un décrochage (fig. <strong>10.</strong>8).<br />
Propagation<br />
asynchrone <strong>de</strong>s OL sur<br />
une section: mixage<br />
Figure <strong>10.</strong>8 : La fréquence d'occurrence <strong>de</strong>s on<strong>de</strong>s lentes sur l'intestin. La<br />
fréquence décroît du duodénum (environ 18/min) à l'iléon (environ 12/min). Ce<strong>la</strong><br />
est dû à une diminution progressive, le long <strong>de</strong> l'intestin, <strong>de</strong> <strong>la</strong> fréquence <strong>de</strong><br />
dépo<strong>la</strong>risation spontanée <strong>de</strong>s fibres lisses. Pour un segment donné <strong>de</strong> l'intestin,<br />
les fréquences sont i<strong>de</strong>ntiques car les zones situées en amont (à fréquence plus<br />
élevée) imposent leur rythme aux zones situées en aval. Ce n'est que lorsque <strong>la</strong><br />
différence <strong>de</strong>s fréquences intrinsèques sera trop gran<strong>de</strong> que se fera un<br />
découp<strong>la</strong>ge. Ce<strong>la</strong> explique que <strong>la</strong> fréquence <strong>de</strong>s OL sur l'intestin grêle diminue<br />
"en marches d'escalier" <strong>et</strong> non selon un continuum.<br />
94
<strong>10.</strong>4. Régu<strong>la</strong>tion <strong>de</strong> <strong>la</strong> <strong>motricité</strong> par le système <strong>nerveux</strong>.<br />
La paroi <strong>de</strong> l'intestin est très riche en neurones (l'intestin contient autant <strong>de</strong><br />
neurones que le système <strong>nerveux</strong> central). On reconnaît une innervation<br />
intrinsèque (majoritaire) située en totalité dans <strong>la</strong> paroi <strong>digestive</strong> <strong>et</strong> une<br />
innervation extrinsèque dont les neurones ont les corps cellu<strong>la</strong>ires en <strong>de</strong>hors <strong>de</strong><br />
<strong>la</strong> paroi <strong>digestive</strong>.<br />
Le système <strong>nerveux</strong> intrinsèque (ou entérique) forme <strong>de</strong>s plexus intramuraux. Le<br />
plexus myentérique (ou plexus d'Auerbach) est le plus volumineux. Il est situé<br />
entre les couches muscu<strong>la</strong>ires longitudinales <strong>et</strong> circu<strong>la</strong>ires responsables du<br />
<strong>contrôle</strong> moteur (fig. <strong>10.</strong>9).<br />
Figure <strong>10.</strong>9. : Plexus <strong>nerveux</strong> <strong>de</strong> l'intestin. Le plexus myentérique (ou<br />
d'Auerbach) est situé entre les couches muscu<strong>la</strong>ires longitudinales <strong>et</strong> circu<strong>la</strong>ires.<br />
Le second système est le sous-muqueux (<strong>de</strong> Meissner).<br />
Le plexus sous-muqueux (<strong>de</strong> Meissner) est moins développé <strong>et</strong> il est impliqué<br />
dans les sécrétions gastro-intestinales <strong>et</strong> le débit sanguin local.<br />
Les plexus reçoivent <strong>de</strong>s afférences du système <strong>nerveux</strong> extrinsèque (système<br />
parasympathique <strong>et</strong> sympathique) <strong>et</strong> ils ém<strong>et</strong>tent <strong>de</strong>s axones efférents<br />
primaires qui remontent vers le système <strong>nerveux</strong> central (substances grise<br />
médul<strong>la</strong>ire, ganglion paravertébraux) ce qui est le support <strong>de</strong> <strong>la</strong> viscéro-<br />
sensibilité (mécanique, chimique,…) (fig. <strong>10.</strong>10).<br />
95
Figure <strong>10.</strong>10 : Coup<strong>la</strong>ge <strong>de</strong> l'innervation intrinsèque <strong>et</strong> <strong>de</strong> l'innervation<br />
extrinsèque. Les plexus <strong>de</strong> <strong>la</strong> paroi <strong>digestive</strong> sont contrôlés par l'innervation<br />
extrinsèque <strong>de</strong> type parasympathique <strong>et</strong> orthosympathique.<br />
Le coup<strong>la</strong>ge entre le système <strong>nerveux</strong> intrinsèque <strong>et</strong> les fibres lisses est assuré<br />
par les cellules <strong>de</strong> Cajal. Les neuromédiateurs sont libérés <strong>de</strong>s varicosités<br />
neuronales avec une organisation synaptique dite "en passage" (fig.<strong>10.</strong>11) ce qui<br />
est différent <strong>de</strong> l'organisation vue dans une p<strong>la</strong>que motrice pour les fibres striées.<br />
Les neurones <strong>de</strong> plexus libèrent différents neuromédiateurs : acétylcholine<br />
(neurone cholinergique), sérotonine (neurone sérotoninergique), VIP (vaso-<br />
intestinal pepti<strong>de</strong>) <strong>et</strong>c.<br />
Plexus<br />
Système <strong>nerveux</strong><br />
extrinsèque<br />
Le principal rôle du système <strong>nerveux</strong> entérique est d'assurer <strong>la</strong> coordination <strong>de</strong>s<br />
2 couches muscu<strong>la</strong>ires dans le sens oral-aboral lors <strong>de</strong> <strong>la</strong> contraction<br />
intestinale avec le réflexe péristaltique comme exemple prototype.<br />
Figure <strong>10.</strong>11 : Coup<strong>la</strong>ge entre l'innervation intrinsèque intramurale <strong>et</strong> <strong>la</strong><br />
muscu<strong>la</strong>ture lisse par les cellules <strong>de</strong> Cajal. Les neurotransm<strong>et</strong>teurs diffusent à<br />
partir <strong>de</strong>s varicosités axonales vers les cellules interstitielles <strong>de</strong> Cajal<br />
(organisation synaptique dite en passage)<br />
Axonal varicosities<br />
Cellule<br />
<strong>de</strong> Cajal<br />
Innervation<br />
extrinsèque<br />
Muscle<br />
lisse<br />
96
<strong>10.</strong>4.1. Rôle du système <strong>nerveux</strong> entérique dans <strong>la</strong><br />
contraction péristaltique <strong>de</strong> l'intestin (loi <strong>de</strong> l'intestin).<br />
Le rôle principal <strong>de</strong> l'innervation intrinsèque est d'assurer <strong>la</strong> coordination du<br />
fonctionnement gastro-intestinal dans le sens oral-aboral. Le réflexe<br />
péristaltique en est un exemple type.<br />
En 1899, Bayliss & Starling énoncent un principe connu <strong>de</strong>puis lors sous le nom<br />
<strong>de</strong> <strong>la</strong> loi <strong>de</strong> l'intestin : lorsque l'intestin est soumis à une excitation (mécanique,<br />
chimique ou électrique) on observe une contraction en amont sur 2-3 cm <strong>et</strong><br />
une inhibition en aval sur 6-7 cm (fig. <strong>10.</strong>12 <strong>et</strong> <strong>10.</strong>13).<br />
Figure <strong>10.</strong>12 La contraction élémentaire <strong>de</strong> l'intestin : l'on<strong>de</strong> péristaltique <strong>et</strong> <strong>la</strong><br />
loi <strong>de</strong> l'intestin.<br />
Zone <strong>de</strong><br />
contraction<br />
Zone <strong>de</strong> relâchement<br />
Temps zéro<br />
5 secon<strong>de</strong>s plus tard<br />
Figure <strong>10.</strong>13: Séquences d'évènements muscu<strong>la</strong>ires pour l'on<strong>de</strong><br />
péristaltique. Grâce à <strong>la</strong> contraction <strong>de</strong> <strong>la</strong> longitudinale, le bol avance dans<br />
l'intestin comme le pied dans une chauss<strong>et</strong>te.<br />
Contraction<br />
circu<strong>la</strong>ire<br />
Contraction<br />
longitudinale<br />
Contraction<br />
circu<strong>la</strong>ire<br />
97
L'intervention du système intramural a été démontrée en faisant <strong>de</strong> multiples<br />
incisions dans <strong>la</strong> paroi <strong>de</strong> l'intestin. Dans ces conditions, on interdit le<br />
développement d'une on<strong>de</strong> péristaltique. Pour que <strong>la</strong> propagation puisse se faire<br />
il faut que les rapports anatomiques entre <strong>la</strong> cellule <strong>et</strong> le plexus d'Auerbach soient<br />
respectés. De même, chez le cobaye, les mouvements péristaltiques<br />
n'apparaissent qu'avec <strong>la</strong> maturation <strong>de</strong>s plexus soit au 28 ème jour <strong>de</strong> gestation. Il<br />
en va <strong>de</strong> même pour le fœtus humain où les premiers mouvements se voient vers<br />
<strong>la</strong> sixième semaine.<br />
La figure <strong>10.</strong>14 montre <strong>de</strong> façon simplifiée le rôle du système entérique. Le point<br />
<strong>de</strong> départ est l'étirement <strong>de</strong> <strong>la</strong> paroi <strong>de</strong> l'intestin par l'arrivée d'un bol. C<strong>et</strong><br />
étirement est détecté par <strong>de</strong>s mécanorécepteurs situés dans <strong>la</strong> paroi. Les<br />
neurones sensoriels font synapse avec <strong>de</strong>s interneurones qui vont assurer (i) en<br />
aval <strong>la</strong> contraction <strong>de</strong> <strong>la</strong> couche longitudinale (qui tire <strong>la</strong> paroi vers l'arrière<br />
(comme une chauss<strong>et</strong>te) <strong>et</strong> qui relâche <strong>la</strong> couche circu<strong>la</strong>ire sous-jacente), (ii) en<br />
amont une contraction <strong>de</strong> <strong>la</strong> couche circu<strong>la</strong>ire (qui pousse le bol vers l'aval <strong>et</strong><br />
un relâchement longitudinal. Les interneurones sont cholinergiques ; les<br />
motoneurones inhibiteurs libèrent du VIP <strong>et</strong> du NO ; les excitateurs libèrent <strong>de</strong><br />
l'acétylcholine <strong>et</strong> <strong>de</strong> <strong>la</strong> substance P.<br />
Figure <strong>10.</strong>14 : Rôle du système entérique dans <strong>la</strong> progression <strong>de</strong> l'on<strong>de</strong><br />
péristaltique. Le point <strong>de</strong> départ est un étirement <strong>de</strong> <strong>la</strong> paroi (str<strong>et</strong>ch) par le<br />
passage du bol. Ce<strong>la</strong> active à <strong>la</strong> fois <strong>de</strong>s neurones inhibiteurs (ce qui va relâcher<br />
les zones plus avales) <strong>et</strong> <strong>de</strong>s neurones stimu<strong>la</strong>teurs (ce qui va entraîner <strong>la</strong><br />
contraction <strong>de</strong> <strong>la</strong> couche circu<strong>la</strong>ire).<br />
Contracted<br />
Re<strong>la</strong>xed<br />
98
<strong>10.</strong>5. L'innervation extrinsèque<br />
L'innervation extrinsèque est double : parasympathique (PS) <strong>et</strong> sympathique<br />
(S) (fig.<strong>10.</strong>15).<br />
Figure <strong>10.</strong>15 : Anatomie générale <strong>de</strong> système <strong>nerveux</strong> autonome<br />
Sympathique Parasympathique<br />
Le PS a pour origine le bulbe avec <strong>la</strong> voie vagale <strong>et</strong> <strong>la</strong> moelle sacrée avec les<br />
nerfs pelviens. L'innervation vagale est surtout importante au niveau <strong>de</strong><br />
l'estomac <strong>et</strong> <strong>de</strong> <strong>la</strong> partie proximale <strong>de</strong> l'intestin. Les fibres PS font synapse au<br />
niveau <strong>de</strong>s plexus (fig. <strong>10.</strong>16) aussi bien sur <strong>la</strong> popu<strong>la</strong>tion <strong>de</strong>s motoneurones<br />
excitateurs cholinergiques que sur les motoneurones inhibiteurs (VIP/NO).<br />
L'innervation vagale est impliquée aussi bien dans les contractions que dans les<br />
relâchements <strong>de</strong> <strong>la</strong> paroi <strong>digestive</strong> (re<strong>la</strong>xation vagale <strong>de</strong> l'estomac liée au réflexe<br />
<strong>de</strong> déglutition). L'innervation vagale a les rôles suivants :<br />
1) <strong>la</strong> re<strong>la</strong>xation <strong>de</strong> l'estomac (zone fundique) pour <strong>la</strong> réception <strong>de</strong>s aliments<br />
2) le tonus vagal diminue <strong>la</strong> fréquence <strong>de</strong>s contractions <strong>de</strong> l'estomac (eff<strong>et</strong><br />
chronotrope négatif) mais en renforce les contractions.<br />
99
3) L'innervation vagale est impliquée dans <strong>la</strong> sécrétion <strong>de</strong> l'estomac (via <strong>la</strong><br />
gastrine)<br />
Figure <strong>10.</strong>16: Contrôle du système <strong>nerveux</strong> intrinsèque par le système <strong>nerveux</strong><br />
extrinsèque (parasympathique <strong>et</strong> sympathique).<br />
<strong>10.</strong>5.1. Le système <strong>nerveux</strong> sympathique<br />
La plupart <strong>de</strong>s fibres sympathiques post-ganglionnaires proviennent <strong>de</strong>s ganglions<br />
coeliaque <strong>et</strong> mésentérique. Les fibres sympathiques sont inhibitrices <strong>et</strong> elles<br />
sont <strong>la</strong>rgement impliquées dans <strong>de</strong> nombreux réflexes d'inhibition à point <strong>de</strong> départ<br />
digestif (iléus paralytique) ou non digestif (péritoine, peau…). Les fibres<br />
sympathiques (avec <strong>la</strong> noradrénaline comme médiateur), agissent en inhibant <strong>de</strong><br />
façon présynaptique les fibres post-ganglionnaires du PS (récepteur <strong>de</strong> type α) ou<br />
par une inhibition directe (récepteur β). Au niveau <strong>de</strong>s sphincters, le système<br />
sympathique joue un rôle excitateur avec <strong>de</strong>s contractions.<br />
<strong>10.</strong>5.2. Les fibres efférentes sensitives<br />
Le tube digestif possè<strong>de</strong> <strong>de</strong> nombreux neurones assurant <strong>la</strong> viscéro-sensibilité<br />
(80% <strong>de</strong>s fibres vagales sont <strong>de</strong>s fibres sensitives). Elles remontent vers le système<br />
<strong>nerveux</strong> central via le nerf sp<strong>la</strong>nchnique.<br />
100
<strong>10.</strong>5.3. Mise en jeu <strong>de</strong> l'innervation extrinsèque : les<br />
réflexes intestinaux longs<br />
L'innervation extrinsèque (sympathique) implique <strong>de</strong> nombreux réflexes:<br />
La distension <strong>de</strong> l'iléon ou l'instil<strong>la</strong>tion d'une solution glucidique dans l'iléon entraîne<br />
une inhibition <strong>de</strong> <strong>la</strong> vidange gastrique <strong>et</strong> <strong>de</strong> <strong>la</strong> <strong>motricité</strong> intestinale. On parle <strong>de</strong> frein<br />
iléal. Il assure le bon déroulement temporel <strong>de</strong> <strong>la</strong> digestion.<br />
La distension d'un segment <strong>de</strong> l'intestin par <strong>de</strong>s gaz ou les manipu<strong>la</strong>tions<br />
chirurgicales dont <strong>la</strong> simple ouverture du péritoine entraîne une inhibition <strong>de</strong> <strong>la</strong><br />
<strong>motricité</strong> intestinale qui passe par les sp<strong>la</strong>nchniques. L'iléus paralytique peut être à<br />
l'origine d'occlusion intestinale réflexe <strong>et</strong> fonctionnelle. C'est ainsi que le<br />
péristaltisme intestinal s'arrête au contact d'un foyer infectieux (appendicite,<br />
péritonite). Certains médicaments ont pour eff<strong>et</strong> <strong>de</strong>s iléus paralytiques (k<strong>et</strong>amine).<br />
L'iléus post-opératoire <strong>de</strong>vient pathologique au-<strong>de</strong>là <strong>de</strong> 48-72h. La reprise <strong>de</strong> <strong>la</strong><br />
<strong>motricité</strong> intestinale doit se faire en quelques heures pour le grêle <strong>et</strong> en 24-48h pour<br />
le côlon. Les morphiniques ralentissent <strong>la</strong> reprise <strong>de</strong> <strong>la</strong> <strong>motricité</strong> <strong>digestive</strong>. Il n'existe<br />
pas <strong>de</strong> traitement préventif <strong>de</strong>s iléus paralytiques (les prokinétiques comme le<br />
métocloprami<strong>de</strong> ou <strong>la</strong> dompéridone – Motilium®, n'ont pas démontré leur efficacité).<br />
Ce réflexe est dû à <strong>la</strong> distension <strong>de</strong> l'estomac (liée à <strong>la</strong> prise <strong>de</strong> nourriture), il peut<br />
déclencher <strong>de</strong>s mouvements <strong>de</strong> masse au niveau du côlon <strong>et</strong> le besoin <strong>de</strong> défécation<br />
(bébé).<br />
101
102