controllo del ciclo cellulare - Bgbunict.it
controllo del ciclo cellulare - Bgbunict.it
controllo del ciclo cellulare - Bgbunict.it

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
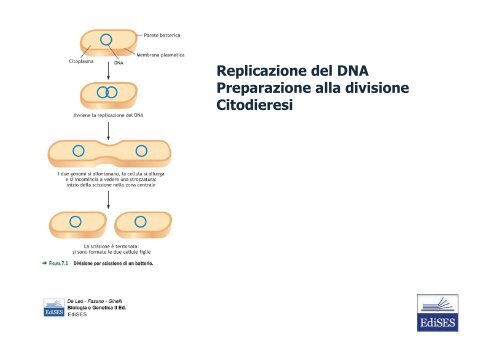
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Replicazione <strong>del</strong> DNA<br />
Preparazione alla divisione<br />
C<strong>it</strong>odieresi
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
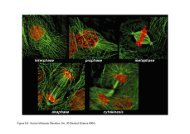
Durata <strong>del</strong> <strong>ciclo</strong> <strong>cellulare</strong>: 24 ore nei fibroblasti; 2<br />
ore nel liev<strong>it</strong>o; 30 minuti nei procarioti
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Quando una cellula non<br />
si divide per un periodo<br />
di tempo lungo, esce dal<br />
<strong>ciclo</strong> <strong>cellulare</strong> e va in<br />
fase G0: neuroni e<br />
cellule muscolari<br />
stazionano per tutta la<br />
loro v<strong>it</strong>a in fase G0,<br />
mentre altri tipi cellulari<br />
restano per lunghi<br />
periodi in G0 per poi<br />
rientrare nel <strong>ciclo</strong> (es,<br />
epatoc<strong>it</strong>i e fibroblasti)<br />
Fase G1: integrazione dei segnali proveniente dall’esterno, es: fattori di cresc<strong>it</strong>a<br />
Fase G2: integrazione dei segnali provenienti dall’interno <strong>del</strong>la cellula, es: verifica<br />
<strong>del</strong>l’integr<strong>it</strong>à <strong>del</strong> genoma<br />
La durata di queste due fasi è diversa nei vari tipi cellulari, tipicamente 9-11 ore G1 e 4<br />
ore per G2; La fase M (m<strong>it</strong>osi) dura circa 1 ora; la fase S dura 8-10 ore.<br />
Durante lo sviluppo embrionale il <strong>ciclo</strong> è molto rapido e G1 e G2 diventano quasi<br />
indistinguibili.
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
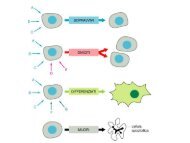
Nei tessuti che rinnovano di continuo<br />
si osserva una rigenerazione<br />
continua per tutta la v<strong>it</strong>a<br />
<strong>del</strong>l’organismo. Da un lato le cellule<br />
si dividono mentre dall’altro si<br />
differenziano e muoiono<br />
Nelle cellule <strong>del</strong>l’epidermide le cellule a contatto con la lamina basale di dividono. Una<br />
<strong>del</strong>le due cellule figlie si differenzia, mentre l’altra rimane staminale (quella che rimane<br />
in contatto con la lamina)
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Il <strong>ciclo</strong> <strong>cellulare</strong> può essere descr<strong>it</strong>to come un’alternanza dei cambiamenti <strong>del</strong>la ploidia.<br />
In G1 sono diploidi; in fase S la s<strong>it</strong>uazione è eterogenea; in G2 avranno il patrimonio<br />
cromosomico raddoppiato (4n)
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
1) Checkpoint che controlla l’ingresso in fase S (transizione G1>S): controlla che il<br />
DNA sia integro e che vi siano gli elementi nutrivi necessari per la cresc<strong>it</strong>a <strong>cellulare</strong><br />
(fattori di cresc<strong>it</strong>a). Se il checkpoint non viene superato la cellula esce dal <strong>ciclo</strong> e va<br />
in G0.<br />
2) Checkpoint che controlla l’ingresso in fase M (transizione G2>M): controlla che il<br />
DNA non abbia sub<strong>it</strong>o danni o mutazioni (verifica <strong>del</strong>la corretta dupicazione prima di<br />
entrare in M)<br />
3) Checkpoint che controlla il completamento <strong>del</strong>la fase M (metafase > c<strong>it</strong>odieresi):<br />
controlla la corretta interazione tra fuso m<strong>it</strong>otico e cromosomi ed il loro appropriato<br />
allineamento lungo la piastra metafasica
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
MPF<br />
cdc2<br />
Esperimenti con gli eterocarionti, con gli ooc<strong>it</strong>i di rana e con i mutanti condizionali di<br />
liev<strong>it</strong>o permisero di identificare un fattore regolativo in regolare alcuni importanti<br />
passaggi <strong>del</strong> <strong>ciclo</strong> <strong>cellulare</strong>
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Nel liev<strong>it</strong>o tra i geni Cdc il più importante è Cdc2. I mutanti si bloccavano nella<br />
transizione G1>S e G2>M. Questo gene è una serina treonina chinasi. Nell’uomo esistono<br />
degli ortologhi di Cdc2 che possono “complementare” la mutazione nel liev<strong>it</strong>o. Questo<br />
dimostra che il macchinario <strong>del</strong> <strong>ciclo</strong> <strong>cellulare</strong> è evolutivamente conservato
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Nel riccio di mare lo zigote va incontro a rapide divisioni cellulari. Durante il <strong>ciclo</strong>, due<br />
proteine si accumulano durante l’interfase per poi scomparire durante la m<strong>it</strong>osi<br />
(degradate). Per via di questa oscillazione legata al <strong>ciclo</strong> <strong>cellulare</strong> sono chiamate cicline<br />
(A e B).<br />
MPF è un complesso formato da due subun<strong>it</strong>à: una catal<strong>it</strong>ica (chinasica), omologa di<br />
Cdc2; ed una regolativa richiesta per l’attiv<strong>it</strong>à enzimatica che si accumula durante il<br />
<strong>ciclo</strong> <strong>cellulare</strong>, omologa <strong>del</strong>le cicline
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Il complesso chinasi-ciclina è il fattore che promuove la m<strong>it</strong>osi; senza la ciclina la chinasi<br />
non è attiva. La chinasi è chiamata Cdk (Cyclin dependent kinase).<br />
I complessi ciclina-Cdk che si formano prima <strong>del</strong>la m<strong>it</strong>osi non sono attivi, ma l’attiv<strong>it</strong>à<br />
chinasica si innesca rapidamente al momento <strong>del</strong>la m<strong>it</strong>osi.<br />
L’attiv<strong>it</strong>à di Cdk viene controllata da chinasi.
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Cdk in assenza di ciclina forma una struttura (Tloop)<br />
che autoinibisce il s<strong>it</strong>o catal<strong>it</strong>ico di Cdk.<br />
L’interazione con la ciclina apre il T-loop (preattivazione).<br />
La treonina 161 che si trovava sul T-loop può<br />
essere fosforilata da CAK (CdK activating kinase).<br />
Questa fosforilazione allontana ulteriormente il Tloop<br />
dal s<strong>it</strong>o catal<strong>it</strong>ico e rafforza l’interazione con la<br />
ciclina (piena attivazione).<br />
Altri due aminoacidi sono importanti per la<br />
regolazione di Cdk: treonina14 e tirosina15. Si<br />
trovano vicino il s<strong>it</strong>o catal<strong>it</strong>ico e una loro<br />
fosforilazione ha una azione inib<strong>it</strong>oria.<br />
La fosforilazione di questi residui è ad opera <strong>del</strong>la<br />
chinasi Wee1. La deforilazione di questi residui è ad<br />
opera <strong>del</strong>la fosfatasi Cdc25.<br />
Il dualismo tra Wee1 e Cdc25 è l’ultima verifica<br />
prima <strong>del</strong>l’attivazione di MPF e <strong>del</strong>la transizione<br />
G2>M)
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Il trans<strong>it</strong>o attraverso la m<strong>it</strong>osi si accompagna alla degradazione <strong>del</strong>la ciclina (e il<br />
conseguente spegnimento <strong>del</strong>l’attiv<strong>it</strong>à chinasica di MPF) ad opera <strong>del</strong> sistema<br />
ubiqu<strong>it</strong>ina-proteasoma.<br />
Durante la m<strong>it</strong>osi il complesso multiproteico APC (Anaphase Promoting Complex)<br />
promuove l’ubiqu<strong>it</strong>inazione <strong>del</strong>la ciclina B
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Le proteine target di MPF:<br />
Le lamine nucleari quando fosforilate depolimerizzano e consentono la vescicolazione<br />
<strong>del</strong>l’involucro nucleare.<br />
GM130 <strong>del</strong>l’Apparato di Golgi che provoca la frammentazione<br />
MAP4 controlla la stabil<strong>it</strong>à dei microtubuli e permette la formazione <strong>del</strong> fuso m<strong>it</strong>otico.<br />
Ruolo nella compattazione <strong>del</strong>la cromatina??? La fosforilazione <strong>del</strong>le condensine,<br />
proteine coinvolte nell’organizzazione <strong>del</strong>la struttura <strong>del</strong>la cromatina
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Nei mammiferi esistono diverse cicline e diversi Cdk.<br />
In diverse fasi <strong>del</strong> <strong>ciclo</strong> si hanno diverse combinazioni ciclina / Cdk<br />
Inoltre differenti complessi ciclina / Cdk sono in grado di modulare altri<br />
processi biologici, come il differenziamento e la trascrizione
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Un <strong>controllo</strong>re fondamentale nella transizione<br />
G1>S è cost<strong>it</strong>u<strong>it</strong>o dal fattore di trascrizione E2F<br />
che come eterodimero (E2F-DP1) si lega ai<br />
promori di molti geni necessari per l’ingresso in<br />
fase S (MYC, ciclina E, E2F).<br />
E2F/DP1 può essere associato alla proteina RB.<br />
RB ha una azione inib<strong>it</strong>oria sulla funzione di E2F.<br />
RB recluta HDAC causando un compattamento<br />
<strong>del</strong>la cromatina ed una inibizione <strong>del</strong>la<br />
trascrizione da parte di E2F/DP1.<br />
Quando la cellula deve progredire nel <strong>ciclo</strong>, RB<br />
viene fosforilato da complessi Cdk/cicline<br />
rilasciando così E2F/DP1.<br />
Nella fase G0. RB non è fosforilato.<br />
Mutazioni di RB (retinoblastoma) impediscono il<br />
<strong>controllo</strong> lim<strong>it</strong>ato <strong>del</strong> <strong>ciclo</strong> <strong>cellulare</strong> e<br />
contribuiscono alla trasformazione cancerosa.<br />
RB è anche bersaglio di alcune proteine virali<br />
(adenovirus, papillomavirus) che inducono la<br />
proliferazione <strong>cellulare</strong> attivando E2F/DP1.
De Leo - Fasano - Ginelli – Biologia e La Genetica, rimozione II Ed. dei – Cap<strong>it</strong>olo fattori 7 di cresc<strong>it</strong>a dal<br />
terreno di coltura porta le cellule in fase<br />
G0.<br />
L’aggiunta di fattori di cresc<strong>it</strong>a nel<br />
terreno stimola il rientro <strong>del</strong>le cellule nel<br />
<strong>ciclo</strong>.<br />
L’attivazione dei recettori per i fattori di<br />
cresc<strong>it</strong>a causa una cascata di reazioni<br />
che porta all’ attivazione trascrizionale di<br />
alcuni geni (proto-oncogeni) come myc,<br />
jun e fos, regolatori <strong>del</strong>la sintesi<br />
proteica e proteine <strong>del</strong> c<strong>it</strong>oscheletro.<br />
Queste proteine portano<br />
successivamente alla trascrizione <strong>del</strong>le<br />
cicline D che attivano Cdk4 che<br />
fosforila RB inattivandolo.<br />
Mutazioni o iperespressione <strong>del</strong>la<br />
ciclina D sono state osservate in diversi<br />
tipi di tumore (linfomi e cancro al seno)
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Quando il DNA è danneggiato (es, rottura <strong>del</strong> doppio<br />
filamento di DNA causato da radiazioni ionizzanti) si<br />
attiva la proteina ATM (atassia telangectasia).<br />
ATM (ser-thre kinase) attiva altre chinasi, es: Chk2<br />
(checkpoint kinase 2)<br />
Queste chinasi a loro volta fosforilano, altre proteine<br />
tra cui CDC25 e TP53.<br />
CDC25 è la fosfatasi che attiva MPF. CDC25<br />
fosforilata fuoriesce dal nucleo e la sua attiv<strong>it</strong>à<br />
fosfatasica viene indebol<strong>it</strong>a. CDK non viene così<br />
attivato, causando il blocco <strong>del</strong> <strong>ciclo</strong>.
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
In genere il livello di TP53 nelle cellule è molto basso per via <strong>del</strong>la continua degradazione<br />
da parte <strong>del</strong> proteasoma. TP53 viene ubiquinato dalla E3 ligasi MDM2.<br />
Se TP53 è fosforilato da Chk2 viene allontanato MDM2 e TP53 si accumula rapidamente<br />
nelle cellule
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7
De Leo - Fasano - Ginelli – Biologia e Genetica, II Ed. – Cap<strong>it</strong>olo 7<br />
Uno dei target trascrizionali di TP53 è p21.<br />
P21 si lega ai complessi Cdk/ciclina<br />
inibendoli (fase G1 ed S) ed arrestando il <strong>ciclo</strong><br />
<strong>cellulare</strong> allo scopo di riparare il DNA.<br />
Alcuni fattori extracellulari sono in grado di<br />
bloccare il <strong>ciclo</strong>, es TGF β. Quando legato al suo<br />
recettore attiva una via di trasduzione <strong>del</strong> segnale<br />
che aumenta l’espressione di un inib<strong>it</strong>ore <strong>del</strong><br />
complesso Cdk4/ciclinaD (CKI) detto p15 (INK4).<br />
Questo causa l’ingresso <strong>del</strong>la cellula in G0 (RB<br />
non può più essere fosforilato). Mutazioni di<br />
questo gene sono state individuate nel<br />
melanoma<br />
L’aumento <strong>del</strong>la dens<strong>it</strong>à <strong>cellulare</strong> provoca un<br />
arresto <strong>del</strong> <strong>ciclo</strong> (inibizione da contatto). In questo<br />
caso viene espresso un altro CKI detto p27.<br />
In genere tutti i CKI, avendo un dominio con una<br />
struttura simile all’ATP, si legano nel s<strong>it</strong>o di Cdk<br />
che dovrebbe legare l’ATP, pregiudicandone<br />
così la funzione.





![Glucidi-Lipidi-010210 [modalità compatibilità] - Bgbunict.it](https://img.yumpu.com/41100335/1/190x134/glucidi-lipidi-010210-modalita-compatibilita-bgbunictit.jpg?quality=85)