

The Plant Vascular System: Evolution, Development and FunctionsF
The Plant Vascular System: Evolution, Development and FunctionsF
The Plant Vascular System: Evolution, Development and FunctionsF

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
Figure 15. Inter-vessel pit structure in angiosperm wood.<br />
Upper-right insert shows brightfield image of two overlapping vessels.<br />
<strong>The</strong>ir common wall is studded with inter-vessel pits as shown in<br />
the main scanning electron microscopy (SEM) image where an intervessel<br />
wall has been sectioned to show individual pit structure. Pits<br />
consist of openings in the secondary wall (apertures) leading to pit<br />
chambers that are spanned by a pit membrane which is the modified<br />
primary cell wall <strong>and</strong> middle lamella of the adjacent vessel elements.<br />
<strong>The</strong> lower-right insert shows an SEM face view of the micro-porous<br />
pit membrane. Scale bars: 5 µm <strong>and</strong> 30 µm for the upper inset.<br />
Micrographs courtesy of Fredrick Lens, Jarmila Pittermann, <strong>and</strong><br />
Brendan Choat.<br />
Inter-conduit pits add substantial flow resistance to the xylem<br />
conduit lumen. An unobstructed lumen conducts water as<br />
efficiently as an ideal cylindrical capillary tube of the same<br />
diameter (Zwieniecki et al. 2001a; Christman <strong>and</strong> Sperry 2010).<br />
<strong>The</strong> most extensive survey indicates that adding inter-conduit<br />
pits increases flow resistivity over that of an unobstructed lumen<br />
by an average factor of 2.8 in conifers with unicellular tracheids<br />
<strong>and</strong> 2.3 in angiosperms with multicellular vessels (Hacke et al.<br />
2006; Pittermann et al. 2006a). <strong>The</strong> lower number for vessels<br />
is not surprising given that they are roughly 10 times longer<br />
than a tracheid of the same diameter (Pittermann et al. 2005),<br />
thereby spacing high resistance pits further apart <strong>and</strong> reducing<br />
the length-normalized resistance (resistivity). What is surprising<br />
is that the greater length of vessels does not have more of<br />
an effect: if inter-vessel pits have the same area-specific pit<br />
resistance as inter-tracheid pits, placing the end-walls 10 times<br />
further apart should increase lumen resistivity by less than a<br />
factor of 1.18. <strong>The</strong> higher observed factor of 2.3 indicates that<br />
inter-vessel pits have higher flow resistance than inter-tracheid<br />
ones, a difference consistent with anatomy <strong>and</strong> estimations<br />
based on modeling (Pittermann et al. 2005).<br />
Inter-vessel pits have nano-porous “homogeneous” pit membranes<br />
(pores usually < 100 nm) (Choat et al. 2008), or<br />
Insights into <strong>Plant</strong> <strong>Vascular</strong> Biology 331<br />
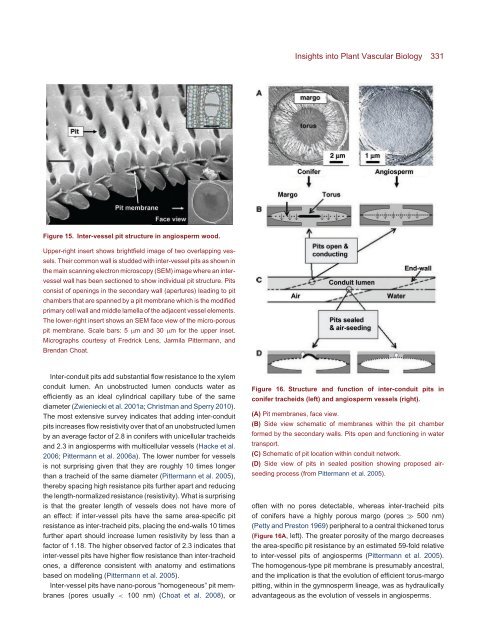
Figure 16. Structure <strong>and</strong> function of inter-conduit pits in<br />
conifer tracheids (left) <strong>and</strong> angiosperm vessels (right).<br />
(A) Pit membranes, face view.<br />
(B) Side view schematic of membranes within the pit chamber<br />
formed by the secondary walls. Pits open <strong>and</strong> functioning in water<br />
transport.<br />
(C) Schematic of pit location within conduit network.<br />
(D) Side view of pits in sealed position showing proposed airseeding<br />
process (from Pittermann et al. 2005).<br />
often with no pores detectable, whereas inter-tracheid pits<br />
of conifers have a highly porous margo (pores ≫ 500 nm)<br />
(Petty <strong>and</strong> Preston 1969) peripheral to a central thickened torus<br />
(Figure 16A, left). <strong>The</strong> greater porosity of the margo decreases<br />
the area-specific pit resistance by an estimated 59-fold relative<br />
to inter-vessel pits of angiosperms (Pittermann et al. 2005).<br />
<strong>The</strong> homogenous-type pit membrane is presumably ancestral,<br />
<strong>and</strong> the implication is that the evolution of efficient torus-margo<br />
pitting, within in the gymnosperm lineage, was as hydraulically<br />
advantageous as the evolution of vessels in angiosperms.













