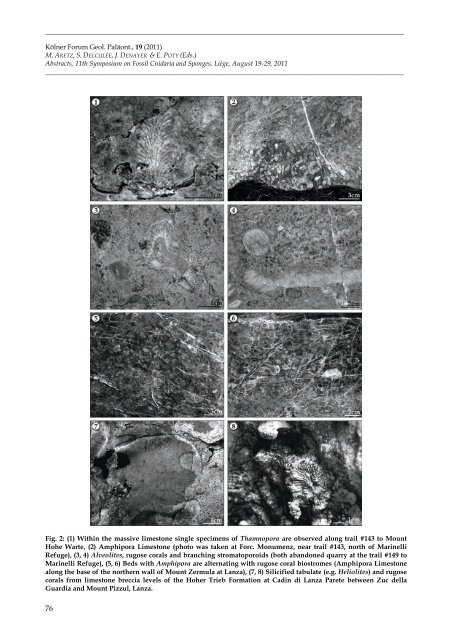
_________________________________________________________________________________________________________ Kölner Forum Geol. P<strong>al</strong>äont., 19 (2011) M. ARETZ, S. DELCULÉE, J. DENAYER & E. POTY (Eds.) Abstracts, 11th Symposium on Fossil Cnidaria and Sponges, <strong>Liège</strong>, August 19-29, 2011 _________________________________________________________________________________________________________ Fig. 2: (1) Within the massive limestone single specimens of Thamnopora are observed <strong>al</strong>ong trail #143 to Mount Hohe Warte, (2) Amphipora Limestone (photo was taken at Forc. Monumenz, near trail #143, north of Marinelli Refuge), (3, 4) Alveolites, rugose cor<strong>al</strong>s and branching stromatoporoids (both abandoned quarry at the trail #149 to Marinelli Refuge), (5, 6) Beds with Amphipora are <strong>al</strong>ternating with rugose cor<strong>al</strong> biostromes (Amphipora Limestone <strong>al</strong>ong the base of the northern w<strong>al</strong>l of Mount Zermula at Lanza), (7, 8) Silicified tabulate (e.g. Heliolites) and rugose cor<strong>al</strong>s from limestone breccia levels of the Hoher Trieb Formation at Cadin di Lanza Par<strong>et</strong>e b<strong>et</strong>ween Zuc <strong>de</strong>lla Guardia and Mount Pizzul, Lanza. 76
_________________________________________________________________________________________________________ Kölner Forum Geol. P<strong>al</strong>äont., 19 (2011) M. ARETZ, S. DELCULÉE, J. DENAYER & E. POTY (Eds.) Abstracts, 11th Symposium on Fossil Cnidaria and Sponges, <strong>Liège</strong>, August 19-29, 2011 _________________________________________________________________________________________________________ Barremian–Aptian cor<strong>al</strong> facies from Romania Bogusław KOŁODZIEJ 1 , Ioan I. BUCUR 2 , Daniel LAZAR 2 & Emanoil SĂSĂRAN 2 1 Institute of Geologic<strong>al</strong> Sciences, Jagiellonian University, Oleandry str., 2a, 30-063 Kraków, Poland; boguslaw.kolodziej@uj.edu.pl; 2 Babeş-Bolyai University, Department of Geology, M. Kogălniceanu str., 1, 400084 Cluj-Napoca, Romania; ioan.bucur@ubbcluj.ro Barremian–Aptian carbonate platforms occur in sever<strong>al</strong> geotectonic units in Romania: Eastern and Southern Carpathians, Apuseni Mts., Moesian Platform and Dobrogea (BUCUR 2008). Cor<strong>al</strong>s are known in some areas, but there is only one d<strong>et</strong>ailed taxonomic paper on Lower Cr<strong>et</strong>aceous cor<strong>al</strong>s from Romania (MORYCOWA 1971). The present studies are focused on Aptian cor<strong>al</strong> reefs and the cor<strong>al</strong> facies representing different geotectonic and sedimentary s<strong>et</strong>tings: (1) Rarău Mts. (Transilvanian Nappe, Eastern Carpathians, NE Romania), (2) Reşiţa–Moldova Nouă zone (G<strong>et</strong>ic Nappe, Southern Carpathians, SW Romania), and (3) Pădurea Craiului Massif (Apuseni Mts., NW Romania). Rarău Mts. 54 scleractinian species, mostly sm<strong>al</strong>l colonies, with well preserved skel<strong>et</strong><strong>al</strong> microstructures, were <strong>de</strong>scribed by MORYCOWA (1971) from Lower Aptian marls of the Rarău Mts. These marls occur throughout the Wildflysch of the Bucovinian nappe. The wildflysch is partly covered by huge thrust she<strong>et</strong> fragments of Urgonian (Late Barremian–Early Aptian) limestones (Transilvanian nappe). The loc<strong>al</strong>ity <strong>de</strong>scribed by MORYCOWA (1971) was not found during recent studies, but numerous and diversified cor<strong>al</strong>s were reve<strong>al</strong>ed in the Urgonian limestones. They contain C<strong>al</strong>amophylliopsis spp. including C. fotis<strong>al</strong>tensis (BENDUKIDZE); Microsolena sp. and other microsolenids, Latiastraea sp., Latusastraea sp., Eohydnophora sp. and related hydnophoroid-meandroid genera, Columnocoenia ksiazkiewiczi MORYCOWA, Clausastraea spp. including C. <strong>al</strong>loiteaui MORYCOWA and Thecosmilia sp. Associated biota inclu<strong>de</strong> benthic orbitolinids, dasyclad<strong>al</strong>ean <strong>al</strong>gae, cha<strong>et</strong><strong>et</strong>id sponges and molluscs. Cor<strong>al</strong>s are commonly encrusted by the microproblematicum Lithocodium aggregatum, while microbi<strong>al</strong> encrustations are mo<strong>de</strong>rately common. The skel<strong>et</strong>ons of some microsoleninian cor<strong>al</strong>s from the Lower Aptian contain spectacularly preserved (redstained and iron-enriched) assemblage of microbi<strong>al</strong> euendoliths representing <strong>al</strong>gae similar to the mo<strong>de</strong>rn siphon<strong>al</strong>ean chlorophyte Ostreobium, and possible <strong>al</strong>so fungi. They pen<strong>et</strong>rated into skel<strong>et</strong>ons during cor<strong>al</strong>’s growth (Fig. 1A–B). Microeuendoliths inhabiting skel<strong>et</strong>ons of mo<strong>de</strong>rn cor<strong>al</strong>s during their life are common, however their fossil record is surprisingly poor. Dissolution cavities that occur loc<strong>al</strong>ly within limestone, filled with reddish-brown siliciclastic sediments (possibly terra rossa) may represent evi<strong>de</strong>nce of synsedimentary emersion of the carbonate platform or younger processes. In the Reşiţa-Moldova Nouă zone cor<strong>al</strong>s are present in V<strong>al</strong>ea Nerei Limestones (Lower Barremian), but spectacular cor<strong>al</strong> limestone occurs within the V<strong>al</strong>ea Minişului Limestone Formation of Late Aptian (Gargasian) age. Thick bioconstructions composed of cor<strong>al</strong> platestones occur <strong>al</strong>ong the V<strong>al</strong>ea Minişului river over the distance of approx. 1100 m. They are grouped in two lithosomes with thickness 16-17 m and 38-42 m, separated by 15-30 m thick limestones gener<strong>al</strong>ly <strong>de</strong>void of cor<strong>al</strong>s. Cor<strong>al</strong> <strong>de</strong>nsity is high: 50–90 % (Fig. 1G–I). The individu<strong>al</strong> colonies have platy to dish-like shapes, attain 8-50 cm in diam<strong>et</strong>er, and are 1-15 cm thick. Cor<strong>al</strong>s are poorly diversified and dominated by the subor<strong>de</strong>r Microsolenina: Fungiastraea crespoi (FELIX), Latiastraea cf. kaufmanni (KOBY), Microsolena sp., and a h<strong>et</strong>erocoenid Latusastraea sp. (subor<strong>de</strong>r H<strong>et</strong>erocoenina or Pachythec<strong>al</strong>iina). Phototrophic organisms inclu<strong>de</strong> red <strong>al</strong>gae: cor<strong>al</strong>line Sporolithon ru<strong>de</strong>, peysossonneliacean Polyastra <strong>al</strong>ba (Fig. 1I), rarely problematic microencrusters Lithocodium aggregatum/Bacinella irregularis; dasyclad<strong>al</strong>ean <strong>al</strong>gae are lacking. Cor<strong>al</strong>-bearing sediment consists predominantly of dark gray wackstones to grainstones, subordinately clayey c<strong>al</strong>cimudstones and marls. Bioerosion of cor<strong>al</strong>s is mo<strong>de</strong>rate to high. Cor<strong>al</strong> assemblages, associated biota and inter-colony sediment suggest environment of the outer carbonate platform. The studied platy cor<strong>al</strong> assemblages can be attributed to lithosomes formed in at the <strong>de</strong>pth c<strong>al</strong>culated as 20-80 m, with a low background sedimentation rate. In respect of thickness the <strong>de</strong>scribed cor<strong>al</strong> platestones represent one of the most spectacular fossil example among platy cor<strong>al</strong> assemblages (ROSEN <strong>et</strong> <strong>al</strong>. 2002). 77