The Frontal lobes - Mahidol University
The Frontal lobes - Mahidol University
The Frontal lobes - Mahidol University

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
19/07/54<br />
415703 Cognitive Neuropsychology<br />
Week 6:<br />
<strong>The</strong> <strong>Frontal</strong> <strong>lobes</strong><br />
Naiphinich Kotchabhakdi, Ph.D.<br />
Director, Salaya Stem Cell R & D Project,<br />
Research Center for Neuroscience,<br />
Institute of Molecular Biosciences,<br />
<strong>Mahidol</strong> <strong>University</strong> Salaya Campus,<br />
999 Phutthamonthol 4 Road, Salaya, Phutthamonthol,<br />
Nakornpathom 73170 Thailand<br />
Email: scnkc@mahidol.ac.th or naiphinich@gmail.com<br />
Web: www.neuroscience.mahidol.ac.th<br />
Main Objectives:<br />
1. <strong>The</strong> <strong>Frontal</strong> <strong>lobes</strong> and their functions<br />
2. <strong>The</strong> Motor System<br />
3. Motor Cortical Organization in the <strong>Frontal</strong> Lobe<br />
4. <strong>The</strong> Prefrontal cortex<br />
5. <strong>The</strong> <strong>Frontal</strong> <strong>lobes</strong> and higher or executive brain<br />
functions<br />
6. Deep brain structures in <strong>Frontal</strong> lobe, e.g.,<br />
Limbic brain structures and their functions<br />
7. Neuropsychology of the <strong>Frontal</strong> <strong>lobes</strong> and<br />
executive brain function disorders.<br />
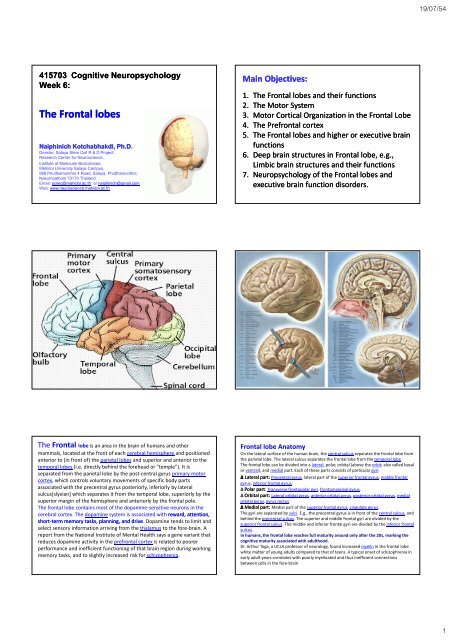
<strong>The</strong> <strong>Frontal</strong> lobe is an area in the brain of humans and other<br />
mammals, located at the front of each cerebral hemisphere and positioned<br />
anterior to (in front of) the parietal <strong>lobes</strong> and superior and anterior to the<br />
temporal <strong>lobes</strong> (i.e. directly behind the forehead or "temple"). It is<br />
separated from the parietal lobe by the post‐central gyrus primary motor<br />
cortex, which controls voluntary movements of specific body parts<br />
associated with the precentral gyrus posteriorly, inferiorly by lateral<br />
sulcus[slyvian] which separates it from the temporal lobe, superiorly by the<br />
superior margin of the hemisphere and anteriorly by the frontal pole.<br />
<strong>The</strong> frontal lobe contains most of the dopamine‐sensitive neurons in the<br />
cerebral cortex. <strong>The</strong> dopamine system is associated with reward, attention,<br />
short‐term term memory tasks, planning, and drive. Dopamine tends to limit and<br />
select sensory information arriving from the thalamus to the fore‐brain. A<br />
report from the National Institute of Mental Health says a gene variant that<br />
reduces dopamine activity in the prefrontal cortex is related to poorer<br />
performance and inefficient functioning of that brain region during working<br />
memory tasks, and to slightly increased risk for schizophrenia.<br />
<strong>Frontal</strong> lobe Anatomy<br />
On the lateral surface of the human brain, the central sulcus separates the frontal lobe from<br />
the parietal lobe. <strong>The</strong> lateral sulcus separates the frontal lobe from the temporal lobe.<br />
<strong>The</strong> frontal lobe can be divided into a lateral, polar, orbital (above the orbit; also called basal<br />
or ventral), and medial part. Each of these parts consists of particular gyri:<br />
∆ Lateral part: Precentral gyrus, lateral part of the superior frontal gyrus, middle frontal<br />
gyrus, inferior frontal gyrus.<br />
∆ Polar part: Transverse frontopolar gyri, frontomarginal gyrus.<br />
∆ Orbital part: Lateral orbital gyrus, anterior orbital gyrus, posterior orbital gyrus, medial<br />
orbital gyrus, gyrus rectus.<br />
∆ Medial part: Medial part of the superior frontal gyrus, cingulate gyrus.<br />
<strong>The</strong> gyri are separated by sulci. E.g., the precentral gyrus is in front of the central sulcus, and<br />
behind the precentral sulcus. <strong>The</strong> superior and middle frontal gyri are divided by the<br />
superior frontal sulcus. <strong>The</strong> middle and inferior frontal gyri are divided by the inferior frontal<br />
sulcus.<br />
In humans, the frontal lobe reaches full maturity around only after the 20s, marking the<br />
cognitive maturity associated with adulthood.<br />
Dr. Arthur Toga, a UCLA professor of neurology, found increased myelin in the frontal lobe<br />
white matter of young adults compared to that of teens. A typical onset of schizophrenia in<br />
early adult years correlates with poorly myelinated and thus inefficient connections<br />
between cells in the fore‐brain<br />
1
19/07/54<br />
Functional areas in the frontal <strong>lobes</strong><br />
<strong>The</strong> motor system is the part of the central nervous system that is involved with<br />
movement. It consists of the pyramidal and extrapyramidal system.<br />
<strong>The</strong> motor pathway also called pyramidal tract or the corticospinal tract start in the motor<br />
center of the cerebral cortex.<br />
<strong>The</strong>re are upper and lower motor neurons in the corticospinal tract.<br />
<strong>The</strong> motor impulses originates in the Giant pyramidal cells or Betz cells of the motor area i.e.<br />
precentral gyrus of cerebral cortex. <strong>The</strong>se are the upper motor neurons (UMN) of the<br />
corticospinal tract. <strong>The</strong> axons of these cells pass in the depth of the cerebral cortex to the<br />
Corona radiata and then to the Internal Capsule passing through the posterior branch of<br />
internal capsule and continue to descend in the Midbrain and the Medulla Oblongata. In<br />
the lower part of Medulla oblongata 80 to 85% of these fibers decussate (pass to the<br />
opposite side) and descend in the White matter of the Lateral funiculus of the spinal cord<br />
on the opposite side. <strong>The</strong> remaining 15 to 20% pass to the same side. Fibers for the<br />
extremities (limbs) pass 100% to the opposite side. <strong>The</strong> fibers of the corticospinal tract<br />
terminate at different levels in the Anterior horn of the Grey matter of the spinal cord.<br />
Here the Lower Motor Neurons (LMN) of the corticospinal cord are located. Peripheral motor<br />
nerves carry the motor impulses from the anterior horn to the voluntary muscles.<br />
Pyramidal motor system:<br />
Corticospinal tracts<br />
tracts is a collection of<br />
axons that travel between the cerebral cortex<br />
of the brain and the spinal cord.<br />
<strong>The</strong> corticospinal tract mostly contains motor axons.<br />
It actually consists of two separate tracts in the<br />
spinal cord: the lateral corticospinal tract and the<br />
anterior corticospinal tract.<br />
An understanding of these tracts leads to an<br />
understanding of why for the most part, one side of<br />
the body is controlled by the opposite side of the<br />
brain.<br />
<strong>The</strong> corticobulbar tract is also considered to be a<br />
pyramidal tract, though it carries signals to motor<br />
neurons of the cranial nerve nuclei, rather than the<br />
spinal cord.<br />
<strong>The</strong> neurons of the corticospinal tracts are referred<br />
to as pyramidal neurons. <strong>The</strong> name comes from the<br />
shape of the corticospinal tracts, which somewhat<br />
resemble pyramids as they pass through the<br />
medulla.<br />
<strong>The</strong> corticospinal tract is concerned specifically with<br />
discrete voluntary skilled movements, especially of<br />
the distal parts of the limbs. (Sometimes called<br />
"fractionated" movements)<br />
<strong>The</strong> motor pathway<br />
<strong>The</strong> corticospinal tract originates from pyramidal cells in layer V of the<br />
cerebral cortex.<br />
About half of its fibres arise from the primary motor cortex. Other<br />
contributions come from the supplementary motor area, premotor<br />
cortex, somatosensory cortex, parietal lobe, and cingulate gyrus. <strong>The</strong><br />
average fiber diameter is in the region of 10μm; around 3% of fibres are<br />
extra‐large (20μm) and arise from Betz cells, mostly in the leg area of the<br />
primary motor cortex.<br />
Upper motor neurons<br />
Upper motor neurons<br />
<strong>The</strong> neuronal cell bodies in the motor cortex, together with their axons<br />
that travel down through the brain stem and spinal cord are commonly<br />
referred to as upper motor neurons. It should be noted however, that<br />
they do not project to muscles, and thus the term 'motor neuron' is<br />
somewhat misleading.<br />
2
19/07/54<br />
Betz ells<br />
Anatomy of the motor cortex<br />
<strong>The</strong> motor cortex can be divided into four main parts:<br />
∆ the primary motor cortex (or M1, Broadman area #4),<br />
responsible for generating the neural impulses controlling<br />
execution of movement<br />
and the secondary motor cortices, including<br />
∆ the posterior parietal cortex, responsible for transforming<br />
visual information into motor commands<br />
∆ the premotor cortex, (Broadman areas #6, 8, 9,10)<br />
responsible for motor guidance of movement and control of<br />
proximal and trunk muscles of the body<br />
∆ and the supplementary motor area (or SMA), responsible<br />
for planning and coordination of complex movements such<br />
as those requiring two hands.<br />
Other brain regions outside the cortex are also of great<br />
importance to motor function, most notably the cerebellum and<br />
subcortical motor nuclei.<br />
<strong>The</strong> primary motor cortex (or M1) is a brain<br />
region that in humans is located in the posterior portion<br />
of the frontal lobe. It works in association with pre‐motor<br />
areas to plan and execute movements. M1 contains large<br />
neurons known as Betz cells which send long axons down<br />
the spinal cord to synapse onto alpha motor neurons<br />
which connect to the muscles. Pre‐motor areas are<br />
involved in planning actions (in concert with the basal<br />
ganglia) and refining movements based upon sensory<br />
input (this requires the cerebellum).<br />
Location of the Primary Motor Cortex (M1)<br />
<strong>The</strong> human primary motor cortex is located in the dorsal part of the precentral gyrus and the<br />
anterior bank of the central sulcus. <strong>The</strong> precentral gyrus is anterior to the postcentral gyrus from<br />
which it is separated by the central sulcus. Its anterior border is the precentral sulcus, while inferiorly<br />
it borders to the lateral fissure (Sylvian fissure). Medially, it is contiguous with the paracentral lobule.<br />
This area can also be identified by Brodmann area number 4.<br />
Layers of the Primary Motor Cortex (M1)<br />
<strong>The</strong> internal pyramidal layer (layer V) of the precentral cortex contains giant (70‐100 micrometers)<br />
pyramidal neurons (a.k.a. Betz cells), which send long axons to the contralateral motor nuclei of the<br />
cranial nerves and to the lower motor neurons in the ventral horn of the spinal cord. <strong>The</strong>se axons<br />
form the corticospinal tract. <strong>The</strong> Betz cells' along with their long axons are referred to as the upper<br />
motor neuron (UMN).<br />
"Homunculus" " or "Little Man"<br />
<strong>The</strong>re is a broadly somatotopic representation of the different body parts in the primary motor<br />
cortex in an arrangement called a motor homunculus (Latin: little man). <strong>The</strong> leg area is located close<br />
to the midline, and the head and face area located laterally on the convex side of the cerebral<br />
hemisphere (motor homunculus). <strong>The</strong> arm and hand motor area is the largest, and occupies the part<br />
of precentral gyrus, between the leg and face area. In humans, the lateral area of the primary motor<br />
cortex is arranged from top to bottom in areas that correspond to the buttocks, torso, shoulder,<br />
elbow, wrist, fingers, thumb, eyelids, lips and jaw. Interior sections of the motor area folding into the<br />
medial longitudinal fissure correspond with the legs.<br />
<strong>The</strong>se areas are not proportional to their size in the body with the lips, face parts and hands enjoying<br />
particularly large areas. Following amputation or paralysis, motor areas can shift to adopt new parts<br />
of the body<br />
3
19/07/54<br />
<strong>The</strong> Modern Era: Neuropsychology Syndrome<br />
Analysis, fMRI Studies, Clinical Neuropsychology<br />
Tests & Clinical Practice<br />
Two representational areas<br />
In primates, the primary motor cortex is unusual in having in its anterior and posterior<br />
areas two representations of the digits and wrist. ] <strong>The</strong> posterior areas can be activated by<br />
attention without any sensory feedback and has been suggested to be important for<br />
initiation of movements, while the anterior areas is dependent on sensory feedback. It<br />
can also be activated by imaginary finger movements and listening to speech done<br />
without actual movements. This anterior representation area has been suggested to be<br />
important executing movements involving complex sensoriomotor interactions.<br />
Pathway<br />
As the motor axons travel down through the cerebral white matter, they move closer<br />
together and form part of the posterior limb of the internal capsule.<br />
<strong>The</strong>y continue down into the brainstem, where some of them, after crossing over to the<br />
contralateral side, distribute to the cranial nerve motor nuclei. (Note: a few motor fibers<br />
synapse with lower motor neurons on the same side of the brainstem).<br />
After crossing over to the contralateral side in the medulla oblongata (pyramidal<br />
decussation), the axons travel down the spinal cord as the lateral corticospinal tract.<br />
Fibers that do not cross over in the brainstem travel down the separate ventral<br />
corticospinal tract and most of them cross over to the contralateral side in the spinal cord,<br />
shortly before reaching the lower motor neurons.<br />
<strong>The</strong> primary motor cortex receive thalamic input from the Ventral lateral nucleus of the<br />
Thalamus.<br />
Blood supply<br />
Branches of the middle cerebral artery provide most of<br />
the arterial blood supply for the primary motor cortex.<br />
<strong>The</strong> medial aspect (leg areas) is supplied by branches of<br />
the anterior cerebral artery.<br />
Neural input from the thalamus<br />
<strong>The</strong> primary motor cortex receive thalamic input from<br />
the Ventral lateral nucleus of the Thalamus.<br />
Pathology<br />
Lesions of the precentral gyrus result in paralysis of the<br />
contralateral side of the body (facial palsy, arm‐/leg<br />
monoparesis, hemiparesis) ‐ see upper motor neuron.<br />
<strong>The</strong> premotor cortex is an area of motor<br />
cortex lying within the frontal lobe of the brain. It<br />
extends 3mm anterior to the primary motor cortex,<br />
near the Sylvian fissure, before narrowing to<br />
approximately 1mm near the medial longitudinal<br />
fissure, which serves as the posterior border for the<br />
prefrontal cortex. <strong>The</strong> premotor cortex is largely<br />
equivalent to Brodmann area 6. Activity within this<br />
region is critical to the sensory guidance of<br />
movement and control of proximal and trunk muscles<br />
of the body.<br />
4
19/07/54<br />
<strong>The</strong> premotor cortex is dysgranular (a transition between the agranular motor cortex and the granular<br />
eulaminate frontal cortex), which means that there is only a faint granular lamina IV. This corresponds to Brodmann<br />
area 6, with the exception that the medial surface of this area is the site of the accessory motor cortex (also known as<br />
the supplementary motor area, or SMA).<br />
Afferents<br />
Subcortical<br />
Cortical<br />
<strong>The</strong> pre‐motor cortex receives inputs originating from other areas of the cortex, including the inferior and<br />
superior parietal lobules (from which it receives multimodal sensory information) and the frontal cortex<br />
(from which it receives information related to attention and motivation) [1]<br />
Efferents<br />
Subcortical<br />
Cortico‐spinal<br />
<strong>The</strong> axons of the premotor cortex contribute to the pyramidal bundle<br />
Striatum<br />
Axons of the Vth layer contribute to the corticostriate connection ; a massive connection involving almost<br />
all parts of the cortex<br />
Thalamus<br />
<strong>The</strong> premotor cortex sends axons to the motor thalamus. This comprises a part receiving cerebellar axons<br />
( nucleus Ventralis Intermedius Vim or VL), a part receiving pallidal axons (nucleus Ventralis oralis VO) and<br />
a part receiving nigral axons (nucleus Ventralis anterio VA). <strong>The</strong> Vim is separated into two parts one<br />
ventrolateral and one mediodorsal VImM. <strong>The</strong> premotor cortex sends axons electively to VImM and VO<br />
Central complex<br />
Subthalamic nucleus<br />
Cortical<br />
Physiology<br />
Mirror neurons are cells located in the premotor cortex, the part of the brain relevant to the planning, selection and<br />
execution of actions. It is a part of the Cerebral cortex.<br />
<strong>The</strong> supplementary<br />
motor area (SMA) is a part<br />
of the sensorimotor cerebral<br />
cortex (perirolandic, i.e. on each<br />
side of the Rolando or central<br />
sulcus). It was included, on purely<br />
cytoarchitectonic arguments, in<br />
area 6 of Brodmann and the<br />
Vogts. It is located on the medial<br />
face of the hemisphere, just in<br />
front of primary motor cortex. This<br />
is an element that appeared late in<br />
evolution, in monkeys, linked to<br />
the appearance of a true medial<br />
pallidum.<br />
Supplementary Motor Area (SMA)<br />
It has been found that the SMA is likely made up of two anatomically and functionally distinct parts, and was divided into the SMA proper<br />
(or: caudal SMA) and the pre‐SMA (or: rostral SMA). In primates, the SMA proper is analogous to area F3, whereas the pre‐SMA is<br />
analogous to area F6.<br />
In monkeys it is a part of the dysgranular cortex. This means an intermediate differentiation between the more posterior agranular motor<br />
cortex and the more anterior granular eulaminate frontal cortex.<br />
Function of SMA<br />
<strong>The</strong> SMA is implicated in the planning of motor actions and bimanual control. In contrast to the premotor cortex, the SMA has been<br />
implicated in actions that are under internal control, such as the performance of a sequence of movements from memory (as opposed to<br />
movements guided by a visual cue).<br />
Pre‐SMA is involved in acquiring new sequences. <strong>The</strong>re is more activity in these neurons when the sequence is new, compared to when it<br />
has been already learned. In contrast, SMA neurons are more active when performing a sequence already learned than one still being<br />
learned. This suggests that the SMA may be more involved in retrieving the sequence. SMA neurons are more active when the task<br />
requires the arrangement of multiple movements in the correct sequence and correct temporal order. For example, some SMA neurons<br />
"prefer" a specific order of movements to be performed. Other SMA neurons fire more for the preparation of a specific rank order. For<br />
example, a neuron can fire more when a monkey is preparing to initiate the third movement, irrespective of the sequence of the three<br />
movements.<br />
SMA and Pre‐SMA can be distinguished by various physiological techniques that delineate two different areas rostrocaudally. Field and<br />
unitary responses to electrical stimulation of the primary motor cortex were distinct in the caudal part, but minimal or absent in the rostral<br />
part. Intracortical microstimulation readily evoked limb or orofacial movements in the caudal part, but only infrequently in the rostral part.<br />
Neuronal responses to visual stimuli prevailed in the rostral part, but somatosensory responses were rare. <strong>The</strong> opposite was true in the<br />
caudal part. <strong>The</strong> rostral part, roughly corresponding to area 6a beta, was operationally defined as the presupplementary motor area (pre‐<br />
SMA). <strong>The</strong> caudal part was redefined as the SMA proper. Single‐cell activity in the pre‐SMA was quantitatively compared with that in the<br />
SMA proper in relation to a trained motor task. Phasic responses to visual cue signals indicating the direction of forthcoming arm‐reaching<br />
movement were more abundant in the pre‐SMA. Activity changes during the preparatory period, which lasted until the occurrence of the<br />
trigger signal for the reaching movement, were more frequent in the pre‐SMA. Phasic, movement‐related activity was more frequent in the<br />
SMA, and its onset was often time locked to the movement onset. In the pre‐SMA, the occurrences of response time locked to the<br />
movement‐trigger signal were more frequent than in the SMA. Among neurons in both areas, directional selectivity was found in all the<br />
cue, preparatory, and movement‐related responses.<br />
Recent considerations of the diverse activities in which the SMA and pre‐SMA play a role suggest existing theories may not fully capture<br />
the fundamental functions of these regions.<br />
<strong>The</strong> SMA is the most dorsal aspect of BA6 (extending medially towards the cingulate gyrus), the ventral aspect which extends down to the<br />
sylvian fissure is the secondary motor cortex (also BA6)<br />
Decussation and synapses<br />
Some of the neuronal cell bodies in the motor cortex send long axons to the motor<br />
cranial nerve nuclei mainly of the contralateral side of the midbrain (corticomesencephalic<br />
tract), pons (Corticopontine tract), and medulla oblongata (cortico‐bulbar<br />
tract), decussating just before they reach their target nuclei.<br />
<strong>The</strong>se are called geniculate fibers. Many more motor cortex neurons, however, extend<br />
fibers all the way down to the spinal cord (corticospinal tract).<br />
► Most of the corticospinal fibers (about 80%) cross over to the contralateral side in the<br />
medulla oblongata (pyramidal decussation). Those that cross in the medulla oblongata<br />
travel in the lateral corticospinal tract.<br />
► 10% enter the lateral corticospinal tract on the same side.<br />
► <strong>The</strong> remainder of them (10%) cross over at the level that they exit the spinal cord, and<br />
these travel in the anterior corticospinal tract.<br />
Whichever of these two tracts it travels in, a corticospinal axon will synapse with another<br />
neuron in the ventral horn. This ventral horn neuron is considered a second‐order neuron<br />
in this pathway, but is not part of the corticospinal tract itself.<br />
5
19/07/54<br />
From cerebral to motor neurons<br />
<strong>The</strong> motor axons move closer together as they travel down through the cerebral white<br />
matter, and form part of the posterior limb of the internal capsule.<br />
<strong>The</strong> motor fibers continue down into the brainstem. <strong>The</strong> bundle of corticospinal axons is<br />
visible as two column‐like structures ("pyramids") on the ventral surface of medulla<br />
oblongata ‐ this is where the name pyramidal tract comes from.<br />
After the decussation, the axons travel down the spinal cord as the lateral corticospinal tract.<br />
Fibers that do not cross over in the medulla oblongata travel down the separate anterior<br />
corticospinal tract, and most of them cross over to the contralateral side in the spinal cord,<br />
shortly before reaching the lower motor neurons.<br />
Lower motor neurons<br />
In the spinal cord, the axons of the upper motor neuron connect (most of them via<br />
interneurons, but to a lesser extent also via direct synapses) with the lower motor neurons,<br />
located in the ventral horn of the spinal cord.<br />
In the brainstem, the lower motor neurons are located in the motor cranial nerve nuclei<br />
(oculomotor, trochlear, motor nucleus of the trigeminal nerve, abducens, facial, accessory,<br />
hypoglossal). <strong>The</strong> lower motor neuron axons leave the brain stem via motor cranial nerves<br />
and the spinal cord via anterior roots of the spinal nerves respectively, end‐up at the<br />
neuromuscular plate and provide motor innervation for voluntary muscles.<br />
<strong>The</strong> Principles of Motor Controls of Movements:<br />
1. <strong>The</strong> central nervous system (CNS) has to choose the right group<br />
of muscles by selecting specific pathways.<br />
2. <strong>The</strong> CNS must give the right amount of excitatory or inhibitory<br />
inputs (“Command”) to specific motoneuron pools<br />
3. <strong>The</strong> excitatory and inhibitory commands must be regulated<br />
“Spatially” and “Temporally”.<br />
4. <strong>The</strong> CNS must regulate the following parameters:<br />
‐ force<br />
‐displacement (distance)<br />
‐ velocity, acceleration or deceleration<br />
<strong>The</strong> extrapyramidal system is a neural network located in the<br />
brain that is part of the motor system involved in the coordination of movement.<br />
<strong>The</strong> system is called "extrapyramidal" to distinguish it from the tracts of the motor<br />
cortex that reach their targets by traveling through the "pyramids" of the medulla.<br />
<strong>The</strong> pyramidal pathways (corticospinal and some corticobulbar tracts) may directly<br />
innervate motor neurons of the spinal cord or brainstem (anterior (ventral) horn<br />
cells or certain cranial nerve nuclei), whereas the extrapyramidal system centers<br />
around the modulation and regulation (indirect control) of anterior (ventral) horn<br />
cells.<br />
Extrapyramidal tracts are chiefly found in the reticular formation of the pons and<br />
medulla, and target neurons in the spinal cord involved in reflexes, locomotion,<br />
complex movements, and postural control. <strong>The</strong>se tracts are in turn modulated by<br />
various parts of the central nervous system, including the nigrostriatal pathway,<br />
the basal ganglia, the cerebellum, the vestibular nuclei, and different sensory<br />
areas of the cerebral cortex. All of these regulatory components can be considered<br />
part of the extrapyramidal system, in that they modulate motor activity without<br />
directly innervating motor neurons.<br />
6
19/07/54<br />
7
19/07/54<br />
8
19/07/54<br />
9
19/07/54<br />
10
19/07/54<br />
11
19/07/54<br />
Corticospinal tract damage<br />
Damage to the descending motor pathways anywhere<br />
along the trajectory from the cerebral cortex to the lower<br />
end of the spinal cord gives rise to a set of symptoms<br />
called the "upper motor neuron syndrome". A few days<br />
after the injury to the upper motor neurons a pattern of<br />
motor signs and symptoms appears, including spasticity,<br />
the decreased vigor (and increased threshold) of<br />
superficial reflexes, a loss of the ability to perform fine<br />
movements, and an extensor plantar response known as<br />
the Babinski sign. [<br />
12
19/07/54<br />
Basal ganglion and<br />
Extrapyramidal<br />
System<br />
13
19/07/54<br />
14
19/07/54<br />
15
19/07/54<br />
<strong>The</strong> extrapyramidal system can be affected in a number of ways,<br />
which are revealed in a range of extrapyramidal symptoms (EPS),<br />
also known as extrapyramidal side‐effects (EPSE), such as akinesia<br />
(inability to initiate movement) and akathisia (inability to remain<br />
motionless).<br />
Extrapyramidal symptoms (EPS) are various movement disorders<br />
such as acute dystonic reactions, pseudoparkinsonism, or akathisia<br />
suffered as a result of taking dopamine antagonists, usually<br />
antipsychotic (neuroleptic) drugs, which are often used to control<br />
psychosis.<br />
<strong>The</strong> Simpson‐Angus Scale (SAS) and the Barnes Akathisia Rating Scale<br />
(BARS) are used to measure extrapyramidal symptoms<br />
16
19/07/54<br />
<strong>The</strong> frontal eye fields (FEF) is a region located in the premotor cortex,<br />
which is part of the frontal cortex of the primate brain.<br />
Function<br />
<strong>The</strong> cortical area called frontal eye fields (FEF) plays an important role in the control of<br />
visual attention and eye movements. Electrical stimulation in the FEF elicits saccadic eye<br />
movements. <strong>The</strong> FEF have a topographic structure and represents saccade targets in<br />
retinotopic coordinates.<br />
<strong>The</strong> frontal eye field is reported to be activated during the initiation of eye movements,<br />
such as voluntary saccades and pursuit eye movements. <strong>The</strong>re is also evidence that it<br />
plays a role in purely sensory processing and that it belongs to a “fast brain” system<br />
through a superior colliculus – medial dorsal nucleus –FEF ascending pathway.In<br />
humans, its earliest activations in regard to visual stimuli occur at 45 ms with activations<br />
related to changes in visual stimuli within 45–60 ms (these are comparable with<br />
response times in the primary visual cortex). This fast brain pathway also provides<br />
auditory input at even shorter times starting at 24 ms and being affected by auditory<br />
characteristics at 30–60 ms. <strong>The</strong> FEF constitutes together with the supplementary eye<br />
fields (SEF), the intraparietal sulcus (IPS) and the superior colliculus (SC) one of the most<br />
important brain areas involved in the generation and control of eye movements,<br />
particularly in the direction contralateral to the frontal eye fields' location.<br />
Brain: <strong>Frontal</strong> eye fields<br />
<strong>Frontal</strong> eye fields is roughly located<br />
between regions #4, #6, and #8<br />
Brodmann area 8, or BA8, is part of the<br />
frontal cortex in the human brain. Situated just<br />
anterior to the premotor cortex (BA6), it includes the<br />
frontal eye fields (so‐named because they are<br />
believed to play an important role in the control of<br />
eye movements). Damage to this area, by stroke,<br />
trauma or infection, causes tonic deviation of the<br />
eyes towards the side of the injury. This finding<br />
occurs during the first few hours of an acute event<br />
such as cerebrovascular infarct (stroke) or<br />
hemorrhage (bleeding).<br />
Distinctive features (Brodmann‐1905): compared to Brodmann area 6‐<br />
1909, area 8 has a diffuse but clearly present internal granular layer (IV);<br />
sublayer 3b of the external pyramidal layer (III) has densely distributed<br />
medium sized pyramidal cells; the internal pyramidal layer (V) has larger<br />
ganglion cells densely distributed with some granule cells interspersed; the<br />
external granular layer (II) is denser and broader; cell layers are more<br />
distinct; the abundance of cells is somewhat greater.<br />
Other Functions<br />
<strong>The</strong> area is involved in the management of uncertainty. A functional<br />
magnetic resonance imaging study demonstrated that brodmann area 8<br />
activation occurs when test subjects experience uncertainty, and that with<br />
increasing uncertainty there is increasing activation.<br />
An alternative interpretation is that this activation in frontal cortex encodes<br />
hope, a higher‐order expectation positively correlated with uncertainty.<br />
Brain: Brodmann area 8<br />
Supplementary eye fields (SEF) are areas on the<br />
dorsal‐medial surface of frontal lobe of the primate brain<br />
that are involved in planning and control of saccadic eye<br />
movements. <strong>The</strong> SEF was first characterized by John Schlag<br />
and colleagues as an area where low intensity electrical<br />
stimulation can evoke saccades, similar to the more lateral<br />
frontal eye fields. More recently it was shown that SEF<br />
stimulation produces coordinated gaze movements of both<br />
the eyes and head. Neural recordings in the SEF show signals<br />
related to both vision and saccades somewhat like the<br />
frontal eye fields and superior colliculus, but currently most<br />
investigators think that the SEF has a special role in high<br />
level aspects of saccade control, like complex spatial<br />
transformations, learned transformations, and executive<br />
cognitive functions<br />
17
19/07/54<br />
<strong>Frontal</strong> lobe Executive Functions<br />
<strong>The</strong> executive functions of the frontal <strong>lobes</strong> involve the ability<br />
to recognize future consequences resulting from current<br />
actions, to choose between good and bad actions (or better<br />
and best), override and suppress unacceptable social<br />
responses, and determine similarities and differences<br />
between things or events. <strong>The</strong>refore, it is involved in higher<br />
mental functions.<br />
<strong>The</strong> frontal <strong>lobes</strong> also play an important part in retaining<br />
longer term memories which are not task‐based. <strong>The</strong>se are<br />
often memories associated with emotions derived from input<br />
from the brain's limbic system. <strong>The</strong> frontal lobe modifies those<br />
emotions to generally fit socially acceptable norms.<br />
Psychological tests that measure frontal lobe function include<br />
finger tapping, Wisconsin Card Sorting Task, and measures of<br />
verbal and figural fluency.<br />
Comparative Neurobiology:<br />
Evolutionary Consideration<br />
COMPARATIVE BRAIN<br />
18
19/07/54<br />
สมองคน<br />
<strong>Frontal</strong> lobe Evolution<br />
For many years, many scientists thought that the frontal<br />
lobe was disproportionately enlarged in humans<br />
compared to other primates. <strong>The</strong>y thought that this was<br />
an important feature of human evolution and was the<br />
primary reason why human cognition is different from<br />
that of the other primates.<br />
However, this view has been challenged by newer<br />
research. Using magnetic resonance imaging to determine<br />
the volume of the frontal cortex in humans, all extant ape<br />
species and several monkey species, Semendeferi et al.<br />
found that the human frontal cortex was not relatively<br />
larger than the cortex in the other great apes but was<br />
relatively larger than the frontal cortex in the lesser apes<br />
and the monkeys<br />
<strong>The</strong> limbic system is also tightly connected to the<br />
prefrontal cortex.<br />
Some scientists contend that this connection is<br />
related to the pleasure obtained from solving problems. To<br />
cure severe emotional disorders, this connection was<br />
sometimes surgically severed, a procedure of<br />
psychosurgery, called a prefrontal lobotomy. Patients who<br />
underwent this procedure often became passive and<br />
lacked all motivation.<br />
<strong>The</strong>re is circumstantial evidence that the limbic<br />
system also provides a custodial function for the<br />
maintenance of a healthy conscious state of mind.<br />
“Psychosurgery”<br />
19
19/07/54<br />
Psychosurgery<br />
In the early 20th century, a medical treatment for mental illness,<br />
first developed by Portuguese neurologist Egas Moniz, involved<br />
damaging the pathways connecting the frontal lobe to the limbic<br />
system. <strong>Frontal</strong> lobotomy (sometimes called frontal leucotomy)<br />
successfully reduced distress but at the cost of often blunting the<br />
subject's emotions, volition and personality. <strong>The</strong> indiscriminate use<br />
of this psychosurgical procedure, combined with its severe side<br />
effects and dangerous nature, gained it a bad reputation. <strong>The</strong><br />
frontal lobotomy has largely died out as a psychiatric treatment.<br />
More precise psychosurgical procedures are still used, although<br />
rarely. <strong>The</strong>y may include anterior capsulotomy (bilateral thermal<br />
lesions of the anterior limbs of the internal capsule) or the bilateral<br />
cingulotomy (involving lesions of the anterior cingulate gyri) and<br />
might be used to treat otherwise untreatable obsessional disorders<br />
or clinical depression<br />
<strong>The</strong>ories of frontal lobe functions can be differentiated into four categories:<br />
∆ Single‐process theories. Posit "that damage to a single process or system is responsible for a<br />
number of different dysexecutive symptoms” (Burgess, 2003, p. 309).<br />
∆ Multi‐process theories. Propose “that the frontal lobe executive system consists of a number of<br />
components that typically work together in everyday actions [(heterogeneity of function)]“ (Burgess,<br />
2003, p. 310).<br />
∆ Construct‐led theories. Assume “that most if not all frontal functions can be explained by one<br />
construct (homogeneity of function) such as working memory or inhibition” (Stuss, 1999, p. 348; cf.<br />
Burgess & Simons, 2005).<br />
∆ Single‐symptom theories. Suggest that a specific dysexecutive symptom (e.g., confabulation) is<br />
related to the processes and construct of the underlying structures (cf. Burgess & Simons, 2005).<br />
Stuss (1999) suggests a differentiation into two categories according to homogeneity and<br />
heterogeneity of function.<br />
Further theoretical approaches to frontal lobe function include:<br />
∆ Grafman's managerial knowledge units (MKU) / structured event complex (SEC) approach (cf. Wood<br />
& Grafman, 2003)<br />
∆ Miller & Cohen's integrative theory of prefrontal functioning (e.g. Miller & Cohen, 2001)<br />
∆ Rolls's stimulus‐reward approach and Stuss's anterior attentional functions (Burgess & Simons,<br />
2005; Burgess, 2003; Burke, 2007).<br />
It may be highlighted that the theories described above differ in their focus on certain<br />
processes/systems or construct‐lets. Stuss (1999) remarks that the question of homogeneity (single<br />
construct) or heterogeneity (multiple processes/systems) of function “may represent a problem of<br />
semantics and/or incomplete functional analysis rather than an unresolvable dichotomy” (p. 348).<br />
However, further research will show if a unified theory of frontal lobe function that fully accounts for<br />
the diversity of functions will be available.<br />
Damage to the frontal <strong>lobes</strong> can lead to a variety of results:<br />
Case Study<br />
Mr. Phineas Gage<br />
Published in<br />
New England<br />
Journal of Medicine<br />
in 1860<br />
20
19/07/54<br />
Brodmann’s area is a region of the cerebral cortex defined based on<br />
its cytoarchitectonics, or organization of cells<br />
Brodmann areas were originally defined and numbered by the German neurologist<br />
Korbinian Brodmann basedonthecytoarchitecture organisation of neurons he<br />
observed in the cerebral cortex using the Nissl stain. Brodmann published his maps<br />
of cortical areas in humans, monkeys, and other species in 1909, along with many<br />
other findings and observations regarding the general cell types and laminar<br />
organization of the mammalian cortex. (<strong>The</strong> same Brodmann area number in<br />
different species does not necessarily indicate homologous areas.)<br />
A more detailed and verifiable cortical map have since been published by Constantin von<br />
Economo and Georg N. Koskinas which greatly improves the quality of the cytoarchitectonic<br />
classifications.<br />
Many of the areas Brodmann defined based solely on their neuronal organization have since<br />
been correlated closely to diverse cortical functions. For example, Brodmann areas 1, 2 and<br />
3aretheprimary somatosensory cortex; area4istheprimary motor cortex; area17isthe<br />
primary visual cortex; and areas 41 and 42 correspond closely to primary auditory cortex.<br />
Higher order functions of the association cortical areas are also consistently localized to the<br />
same Brodmann areas by neurophysiological, functional imaging, and other methods (e.g.,<br />
the consistent localization of Broca's speech and language area to the left Brodmann areas<br />
44 and 45). However, functional imaging can only identify the approximate localization of<br />
brain activations in terms of Brodmann areas since their actual boundaries in any individual<br />
brain requires its histological examination.<br />
Brodmann areas for human & non‐human primates<br />
Brodmann’s areas 3D<br />
map: Lateral Surface<br />
map: Medial Surface<br />
Brodmann areas for human & non‐human primates<br />
21
19/07/54<br />
22
19/07/54<br />
Paul Pierre Broca (June 28, 1824 –<br />
July 9, 1880) was a French physician,<br />
anatomist, and anthropologist. He was<br />
born in Sainte-Foy-la-Grande, France.<br />
He is best known for his research on<br />
Broca's area, a region of the frontal<br />
lobe that has been named after him.<br />
Broca's legacy<br />
<strong>The</strong> discovery of Broca's area revolutionized the<br />
understanding of speech production. New research<br />
has found that dysfunction in the area may lead to<br />
other speech disorders such as stuttering and<br />
apraxia of speech. Recent anatomical<br />
neuroimaging studies have shown that the pars<br />
opercularis of Broca's area is anatomically smaller<br />
in individuals who stutter whereas the pars<br />
triangularis appears to be normal.<br />
Speech research<br />
Broca is most famous for his discovery of the speech production center of the brain<br />
located in the ventroposterior region of the frontal <strong>lobes</strong> (now known as Broca's<br />
area). He arrived at this discovery by studying the brains of aphasic patients. His first<br />
patient in the Bicêtre Hospital was Leborgne, nicknamed "Tan" due to his inability to<br />
clearly speak any words other than "tan".<br />
In 1861, through post‐mortem autopsy, Broca determined that Tan had a lesion<br />
caused by syphilis in the left cerebral hemisphere. This lesion was determined to<br />
cover the area of the brain important for speech production. (Although history<br />
credits this discovery to Broca, it should be noted that another French neurologist,<br />
Marc acDax, made similar observations o s a generation e earlier.) e ) Today the brains ba sof many<br />
of Broca's aphasic patients are still preserved in the Musée Dupuytren, and his<br />
collection of casts in the Musée d'Anatomie Delmas‐Orfila‐Rouvière.<br />
Patients with damage to Broca's area and/or to neighboring regions of the left<br />
inferior frontal lobe are often categorized clinically as having Broca's aphasia. This<br />
type of aphasia, which often involves impairments in speech output, can be<br />
contrasted with Wernicke's aphasia, named for Karl Wernicke, which is<br />
characterized by damage to more posterior regions of the left hemisphere (in the<br />
superior temporal lobe), and by greater impairments in speech comprehension. This<br />
is an example of a double dissociation, an important tool used by<br />
neuropsychologists to investigate brain function.<br />
23
19/07/54<br />
Paul Broca (1824-1880)<br />
Tan’s Brain<br />
(removed in 1861)<br />
Now in the Museum of Man, Paris<br />
Expressive aphasia, known as Broca's aphasia in clinical<br />
neuropsychology and agrammatic aphasia in cognitive<br />
neuropsychology, is an aphasia caused by damage to or<br />
developmental issues in anterior regions of the brain, including (but<br />
not limited to) the left inferior frontal region known as Broca's area<br />
(Brodmann area 44 and Brodmann area 45)<br />
Presentation<br />
Sufferers of this form of aphasia exhibit the common problem of agrammatism. For them, speech is difficult to initiate,<br />
non‐fluent, labored, and halting. Similarly, writing is difficult as well. Intonation and stress patterns are deficient.<br />
Language g is reduced to disjointed words and sentence construction is poor, omitting function words and inflections<br />
(bound morphemes). A person with expressive aphasia might say "Son ... <strong>University</strong> ... Smart ... Boy ... Good ... Good ... "<br />
For example, in the following passage, a Broca's aphasic patient is trying to explain how he came to the hospital for<br />
dental surgery:<br />
Yes... ah... Monday... er... Dad and Peter H... (his own name), and Dad.... er... hospital... and ah... Wednesday...<br />
Wednesday, nine o'clock... and oh... Thursday... ten o'clock, ah doctors... two... an' doctors... and er... teeth...<br />
yah.[1]<br />
In extreme cases, patients may be only able to produce a single word. <strong>The</strong> most famous case of this was Paul Broca's<br />
patient Leborgne, nicknamed "Tan", after the only syllable he could say. Even in such cases, over-learned and rote-learned<br />
speech patterns may be retained—for instance, some patients can count from one to ten, but cannot produce the same<br />
numbers in ordinary conversation.<br />
While word comprehension is generally preserved, meaning interpretation dependent on syntax and phrase structure is<br />
substantially impaired. This can be demonstrated by using phrases with unusual structures. A typical Broca's aphasic<br />
patient will misinterpret "the dog is bitten by the man" by switching the subject and object. Patients who recover go on to<br />
say that they knew what they wanted to say but could not express themselves. Residual deficits will often be seen.<br />
Broca's area is a region of the hominid brain<br />
with functions linked to speech production.<br />
<strong>The</strong> production of language has been linked to the<br />
Broca’s area since Pierre Paul Broca reported<br />
impairments in two patients. <strong>The</strong>y had lost the<br />
ability to speak after injury to the posterior inferior<br />
frontal gyrus of the brain. Since then, the<br />
approximate region he identified has become<br />
known as Broca’s area, and the deficit in language<br />
production as Broca’s aphasia. Broca’s area is now<br />
typically defined in terms of the pars opercularis<br />
and pars triangularis of the inferior frontal gyrus,<br />
represented in Brodmann’s<br />
cytoarchitectonic map<br />
as areas 44 and 45. Studies of chronic aphasia have<br />
implicated an essential role of Broca’s area in<br />
various speech and language functions. Further,<br />
functional MRI studies have also identified<br />
activation patterns in Broca’s area associated with<br />
various language tasks. However, slow destruction<br />
of the Broca's area by brain tumors can leave<br />
speech relatively intact suggesting its functions can<br />
shift to nearby areas in the brain.<br />
Brodmann area 44, , or BA44<br />
44, is part of the frontal<br />
cortex in the human brain. Situated just anterior to premotor<br />
cortex (BA6) and on the lateral surface, inferior to BA9.<br />
This area is also known as pars opercularis (of the inferior<br />
frontal gyrus), and it refers to a subdivision of the<br />
cytoarchitecturally defined frontal region of cerebral cortex. In<br />
the human it corresponds approximately to the opercular part<br />
of inferior frontal gyrus (H). Thus, it is bounded caudally by the<br />
inferior precentral sulcus (H) and rostrally by the anterior<br />
ascending limb of lateral sulcus (H). It surrounds the diagonal<br />
sulcus (H). In the depth of the lateral sulcus it borders on the<br />
insula. Cytoarchitectonically it is bounded caudally and dorsally<br />
by the agranular frontal area 6, dorsally by the granular frontal<br />
area 9 and rostrally by the triangular area 45 (Brodmann‐1909).<br />
Together with left‐hemisphere BA 45, the left hemisphere . BA<br />
44 comprises Broca's area a region involved in semantic tasks.<br />
Some data suggest that BA44 is more involved in phonological<br />
and syntactic processing. Some recent findings also suggest the<br />
implication of this region in music perception. In 95.5% of righthanders<br />
and 61.4% of left‐handers, therefore about 90% of the<br />
clinical population, speech is lateralised in the left hemisphere.<br />
Brain: Brodmann area 44<br />
7/19/2011 NEUROPSYCHIATRY 143<br />
24
19/07/54<br />
Brodmann area 45 (BA45),<br />
is part of the frontal<br />
cortex in the human brain. Situated on the lateral surface,<br />
inferior to BA9 and adjacent to BA46. This area is also known as<br />
pars triangular (of the inferior frontal gyrus). In the human, it<br />
occupies the triangular part of inferior frontal gyrus (H) and, surrounding<br />
the anterior horizontal limb of lateral sulcus (H), a portion of the orbital<br />
part of inferior frontal gyrus (H). Bounded caudally by the anterior<br />
ascending limb of lateral sulcus (H), it borders on the insula in the depth<br />
of the lateral sulcus. Cytoarchitectonically it is bounded caudally by the<br />
opercular area 44 (BA44), rostrodorsally by the middle frontal area 46<br />
(BA46) and ventrally by the orbital area 47 (BA47) (Brodmann‐1909).<br />
Together with BA 44 it comprises Broca's area, a region which is active in<br />
semantic tasks, such as semantic decision tasks (determining whether a<br />
word represents an abstract or a concrete entity) and generation tasks<br />
(generating a verb associated with a noun).<br />
<strong>The</strong> precise role of BA45 in semantic tasks remains controversial. For<br />
some researchers, its role would be to subserve semantic retrieval or<br />
semantic working memory processes. Under this view, BA44 and BA45<br />
would together guide recovery of semantic information and evaluate the<br />
recovered information with regards to the criterion appropriate to a<br />
given context. A slightly modified account of this view is that activation<br />
of BA45 is needed only under controlled semantic retrieval, when strong<br />
stimulus‐stimulus associations are absent. For other researchers, BA45's<br />
role is not restricted to semantics per se, but to all activities which<br />
require task‐relevant representations from among competing<br />
representations.<br />
Brain: Brodmann area 45<br />
Brodmann area 47, , or BA47<br />
47, is part of<br />
the frontal cortex in the human brain. Curving from the<br />
lateral surface of the frontal lobe into the ventral<br />
(orbital) frontal cortex. It is below areas BA10 and BA45,<br />
and beside BA11.<br />
This area is also known as orbital area 47. In the human,<br />
on the orbital surface it surrounds the caudal portion of<br />
the orbital sulcus (H) from which it extends laterally into<br />
the orbital part of inferior frontal gyrus (H).<br />
Cytoarchitectonically yoac eco cayit is bounded caudally by the<br />
triangular area 45, medially by the prefrontal area 11 of<br />
Brodmann‐1909, and rostrally by the frontopolar area 10<br />
(Brodmann‐1909).<br />
It incorporates the region that Brodmann identified as<br />
"Area 12" in the monkey, and therefore, following the<br />
suggestion of Michael Petrides, some contemporary<br />
neuroscientists refer to the region as "BA47/12."<br />
BA47 has been implicated in the processing of syntax<br />
in spoken and signed languages, and more recently in<br />
musical syntax.<br />
Brain: Brodmann area 47<br />
<strong>The</strong> Wernicke's area is classically located as<br />
the posterior section of the superior temporal gyrus<br />
(STG) in the left (or dominant) cerebral hemisphere.<br />
This area encircles the auditory cortex on the Sylvian<br />
fissure (part of the brain where the temporal lobe<br />
and parietal lobe meet). This area is<br />
neuroanatomically described as the posterior part of<br />
Brodmann area 22.<br />
However, there is an absence of consistent<br />
definitions as to its location. Some identify it with<br />
the unimodal auditory association in the superior<br />
temporal gyrus anterior to the primary auditory<br />
cortex. Others include also adjacent parts of the<br />
heteromodal cortex in BA 39 and BA40 in the<br />
parietal lobe.<br />
While previously thought to connect Wernicke's area<br />
and Broca's area, new research demonstrates that<br />
the arcuate fasciculus instead connects to posterior<br />
receptive areas with premotor/motor areas, and not<br />
to Broca's area<br />
Wernicke and aphasia<br />
Wernicke's area is named after Carl Wernicke, a German neurologist and psychiatrist who, in<br />
1874, hypothesized a link between the left posterior section of the superior temporal gyrus<br />
and the reflexive mimicking of words and their syllables that associated the sensory and motor<br />
images of spoken words. He did this on the basis of the location of brain injuries that caused<br />
aphasia. Receptive aphasia in which such abilities are preserved is now sometimes called<br />
Wernicke's aphasia. In this condition there is a major impairment of language comprehension,<br />
while speech retains a natural‐sounding rhythm and a relatively normal syntax. Language as a<br />
result is largely meaningless (a condition sometimes called fluent or jargon aphasia).<br />
While neuroimaging and lesion evidence generally support the idea that malfunction of or damage to<br />
Wernicke's area is common in people with receptive aphasia, this is not always so. Some people may use<br />
the right hemisphere for language, and isolated damage of Wernicke's area cortex (sparing white matter<br />
and other areas) may not cause severe receptive aphasia. Even when patients with Wernicke's area lesions<br />
have comprehension deficits, these are usually not restricted to language processing alone. For example,<br />
one study found that patients with posterior lesions also had trouble understanding nonverbal sounds like<br />
animal and machine noises. In fact, for Wernicke's area, the impairments in nonverbal sounds were<br />
statistically stronger than for verbal sounds.<br />
Right homologous area<br />
Research using Transcranial magnetic stimulation suggests that the area corresponding to the Wernicke’s<br />
area in the non‐dominant cerebral hemisphere has a role in processing and resolution of subordinate<br />
meanings of ambiguous words—such as (‘‘river’’) when given the ambiguous word (‘‘bank’’). In contrast,<br />
the Wernicke's area in the dominant hemisphere processes dominant word meanings (‘‘teller’’ given<br />
‘‘bank’’).<br />
25
19/07/54<br />
<strong>The</strong> Wernicke-Geschwind model of<br />
language<br />
Wernicke created an early neurological model<br />
of language, that later was revived by Norman<br />
Geschwind. <strong>The</strong> model is known as the<br />
Wernicke‐Geschwind model.<br />
26
19/07/54<br />
Prefrontal<br />
cortex<br />
Brodmann area 9, , or BA9, is part of the<br />
frontal cortex in the human brain. It<br />
contributes to the dorsolateral prefrontal<br />
cortex.<br />
Brodmann area 9 refers to a cytoarchitecturally defined<br />
portion of the frontal lobe of the guenon (Old world<br />
monkeys). Brodmann‐1909 regarded it on the whole as<br />
topographically and cytoarchitecturally homologous to the<br />
granular frontal area 9 and frontopolar area 10 in the<br />
human. Distinctive features (Brodmann‐1905): unlike<br />
Brodmann area 6‐1909, area 9 has a distinct internal<br />
granular layer (IV); unlike Brodmann area 6 or Brodmann<br />
area 8‐1909 its internal pyramdal layer (V) is divisible into<br />
two sublayers, an outer layer 5a of densely distributed<br />
medium sized ganglion cells that partially merges with<br />
layer IV, and an inner, clearer, cell‐poor layer 5b; the<br />
pyramidal cells of sublayer 3b of the external pyramidal<br />
layer (III) are smaller and sparser in distribution; the<br />
external granular layer (II) is narrow, with small numbers<br />
of sparsely distributed granule cells.<br />
Brain: Brodmann area 9<br />
<strong>The</strong> dorsolateral prefrontal cortex (DL‐PFC or DLPFC),<br />
according to a more restricted definition, is roughly equivalent to Brodmann areas 9 and 46.<br />
According to a broader definition DL‐PFC consists of the lateral portions of Brodmann areas<br />
9 – 12, of areas 45, 46, and the superior part of area 47.<strong>The</strong>se regions mainly receive their<br />
blood supply from the middle cerebral artery. With respect to neurotransmitter systems,<br />
there is evidence that dopamine plays a particularly important role in DL‐PFC.<br />
DL‐PFC is connected to the orbitofrontal cortex, and to a variety of brain areas, which<br />
include the thalamus, parts of the basal ganglia (the dorsal caudate nucleus), the<br />
hippocampus, and primary and secondary association areas of neocortex, including<br />
posterior temporal, parietal, and occipital areas.<br />
DL‐PFC is the last area, 45th, to develop myelinate in the human cerebrum<br />
DL‐PFC serves as the highest h cortical area responsible for motor planning, organization, i and<br />
regulation. It plays an important role in the integration of sensory and mnemonic<br />
information and the regulation of intellectual function and action. It is also involved in<br />
working memory. However, DL‐PFC is not exclusively responsible for the executive<br />
functions. All complex mental activity requires the additional cortical and subcortical<br />
circuits with which the DL‐PFC is connected.<br />
Damage to the DL‐PFC can result in the dysexecutive syndrome, [4] which leads to problems<br />
with affect, social judgement, executive memory, abstract thinking and intentionality. [<br />
Lucid dream states<br />
More recent research has found a connection between the DL‐PFC and lucid dream states<br />
in which executive function is retained<br />
27
19/07/54<br />
Brodmann area 10, or BA10<br />
is the<br />
frontopolar part of the frontal cortex in the human brain.<br />
BA10 was originally defined in terms of microscopic<br />
cytoarchitecturic traits in autopsy brains; modern functional<br />
imaging research cannot directly identify these boundaries<br />
and the terms anterior prefrontal, rostral prefrontal cortex<br />
and frontopolar prefrontal cortex are used to refer to the<br />
area in the most anterior part of the frontal cortex that<br />
approximates to or principally covers BA10.<br />
BA10 is the largest cytoarchitectonic area in the human<br />
brain. It has been described as "one of the least well<br />
understood dregions of the human brain". Present research<br />
suggests that it is involved in strategic processes in memory<br />
retrieval and executive function. During human evolution,<br />
the functions in this area resulted in its expansion relative to<br />
the rest of the brain.<br />
Although this region is extensive in humans, its function is poorly understood.<br />
Koechlin & Hyafil have proposed that processing of 'cognitive branching' is the<br />
core function of the frontopolar cortex. Cognitive branching enables a<br />
previously running task to be maintained in a pending state for subsequent<br />
retrieval and execution upon completion of the ongoing one. Many of our<br />
complex behaviors and mental activities require simultaneous engagement of<br />
multiple tasks, and they suggest the anterior prefrontal cortex may perform a<br />
domain‐general function in these scheduling operations. However, other<br />
hypotheses have also been proffered, such as those by Burgess et al.<br />
Brain: Brodmann area 10<br />
Brodmann area 11 is one of Brodmann's<br />
cytologically defined regions of the brain. It is<br />
involved in planning, reasoning, and decision<br />
making.<br />
Brodmann area 11, or BA11, is part of the frontal cortex in the<br />
human brain. BA11 covers the medial part of the ventral surface of<br />
the frontal lobe.<br />
Prefrontal area 11 of Brodmann‐1909 is a subdivision of the frontal<br />
lobe in the human defined on the basis of cytoarchitecture. Defined<br />
and illustrated in Brodmann‐1909, it included the areas<br />
subsequently illustrated in Brodmann‐10 as prefrontal area 11 and<br />
rostral area 12.<br />
prefrontal area 11 is a subdivision of the cytoarchitecturally defined<br />
frontal region of cerebral cortex of the human. As illustrated in<br />
Brodmann‐10, It constitutes most of the orbital gyri, gyrus rectus<br />
and the most rostral portion of the superior frontal gyrus. It is<br />
bounded medially by the inferior rostral sulcus (H) and laterally<br />
approximately by the frontomarginal sulcus (H). Cytoarchitecturally<br />
it is bounded on the rostral and lateral aspects of the hemisphere<br />
by the frontopolar area 10, the orbital area 47, and the triangular<br />
area 45; on the medial surface it is bounded dorsally by the rostral<br />
area 12 and caudally by the subgenual area 25. In an earlier map,<br />
the area labeled 11, i.e., prefrontal area 11 of Brodmann‐1909, was<br />
larger; it included the area now designated rostral area 12.<br />
Brain: Brodmann area 11<br />
Brodmann area 46, or BA46<br />
46, is part of the frontal cortex<br />
in the human brain. It is between BA10 and BA45.<br />
BA46 is known as middle frontal area 46. In the human brain it<br />
occupies approximately the middle third of the middle frontal<br />
gyrus and the most rostral portion of the inferior frontal gyrus.<br />
Brodmann area 46 roughly corresponds with the dorsolateral<br />
prefrontal cortex (DLPFC), although the borders of area 46 are<br />
based on cytoarchitecture rather than function. <strong>The</strong> DLPFC also<br />
encompasses part of granular frontal area 9, directly adjacent on<br />
the dorsal surface of the cortex.<br />
Cytoarchitecturally, BA46 is bounded dorsally by the granular frontal area<br />
9, rostroventrally by the frontopolar area 10 and caudally by the<br />
triangular area 45 (Brodmann‐1909). <strong>The</strong>re is some discrepancy between<br />
the extent of BA8 (Brodmann‐1905) and the same area as described by<br />
Walker (1940)<br />
<strong>The</strong> DLPFC plays a role in sustaining attention and working<br />
memory. Lesions to the DLPFC impair short‐term memory and<br />
cause difficulty inhibiting responses. Lesions may also eliminate<br />
much of the ability to make judgements about what's relevant<br />
and what's not as well as causing problems in organization.<br />
<strong>The</strong> DLPFC has recently been found to be involved in exhibiting<br />
self‐control. <strong>The</strong> dorsolateral prefrontal cortex, which is one of the few<br />
areas deactivated during REM sleep. Neuroscientist J. Allan Hobson has<br />
hypothesized that activation of the dorsolateral prefrontal cortex produce<br />
lucid dreams.<br />
Brain: Brodmann area 46<br />
<strong>The</strong> limbic system is also tightly connected to the<br />
prefrontal cortex.<br />
Some scientists contend that this connection is<br />
related to the pleasure obtained from solving problems. To<br />
cure severe emotional disorders, this connection was<br />
sometimes surgically severed, a procedure of<br />
psychosurgery, called a prefrontal lobotomy. Patients who<br />
underwent this procedure often became passive and<br />
lacked all motivation.<br />
<strong>The</strong>re is circumstantial evidence that the limbic<br />
system also provides a custodial function for the<br />
maintenance of a healthy conscious state of mind.<br />
28
19/07/54<br />
<strong>The</strong> Brodmann area 32<br />
32, also known in the<br />
human brain as the dorsal anterior cingulate area<br />
32, refers to a subdivision of the cytoarchitecturally<br />
defined cingulate region of cerebral cortex. In the<br />
human it forms an outer arc around the anterior<br />
cingulate gyrus. <strong>The</strong> cingulate sulcus defines<br />
approximately its inner boundary and the superior<br />
rostral sulcus (H) its ventral boundary; rostrally it<br />
extends almost to the margin of the frontal lobe.<br />
Cytoarchitecturally it is bounded internally by the<br />
ventral anterior cingulate area 24, externally by<br />
medial margins of the agranular frontal area 6,<br />
intermediate frontal area 8, granular frontal area 9,<br />
frontopolar area 10, and prefrontal area 11‐1909.<br />
(Brodmann19‐09).<br />
Dorsal region of anterior cingulate gyrus is<br />
associated with rational thought processes, most<br />
notably active during the Stroop task.<br />
Brain: Brodmann area 32<br />
Stroop effect is a<br />
demonstration of the reaction time of a<br />
task. When the name of a color (e.g.,<br />
"blue," "green," or "red") is printed in a<br />
color not denoted by the name (e.g., the<br />
word "red" printed in blue ink instead of<br />
red ink), naming the color of the word<br />
takes longer and is more prone to errors<br />
than when the color of the ink matches the<br />
name of the color. <strong>The</strong> effect is named<br />
after John Ridley Stroop who first<br />
published the effect in English in 1935. <strong>The</strong><br />
effect had previously been published in<br />
Germany in 1929. <strong>The</strong> original paper has<br />
been one of the most cited papers in the<br />
history of experimental psychology, leading<br />
to more than 700 replications. <strong>The</strong> effect<br />
has been used to create a psychological<br />
test (Stroop<br />
Test) that is widely used in<br />
clinical practice and investigation.<br />
This test is considered to measure selective<br />
attention, cognitive flexibility and processing<br />
speed, and it is used as a tool in the<br />
evaluation of executive functions. An<br />
increased interference effect is found in<br />
disorders such as brain damage, dementias<br />
and other neurodegenerative diseases,<br />
attention‐deficit hyperactivity disorder, or a<br />
variety of mental disorders such as<br />
schizophrenia, addictions, and depression<br />
<strong>The</strong> anterior cingulate cortex has been related<br />
to the processing of the Stroop effect<br />
Figure 1 from Experiment 2 of the original description of the Stroop<br />
Effect (1935). 1 is the time that it takes to name the color of the dots<br />
while 2 is the time that it takes to say the color when there is a conflict<br />
with the written word<br />
Brodmann area 24 is part of the anterior<br />
cingulate in the human brain.<br />
In the human this area is known as ventral anterior<br />
cingulate area 24, and it refers to a subdivision of the<br />
cytoarchitecturally defined cingulate cortex region of<br />
cerebral cortex (area cingularis anterior ventralis). It<br />
occupies most of the anterior cingulate gyrus in an arc<br />
around the genu of corpus callosum. Its outer border<br />
corresponds approximately to the cingulate sulcus.<br />
Cytoarchitecturally it is bounded internally by the<br />
pregenual area 33, externally by the dorsal anterior<br />
cingulate area 32, and caudally by the ventral<br />
posterior cingulate area 23 and the dorsal posterior<br />
cingulate area 31.<br />
Francis Crick, one of the discoverers of DNA, listed<br />
area 24 as the seat of free will because of its<br />
centrality in abulia and amotivational syndromes.<br />
Brain: Brodmann area 24<br />
Aboulia or Abulia (from the Greek "αβουλία", meaning<br />
"non‐will"), in neurology, refers to a lack of will or initiative<br />
and is one of the Disorders of Diminished Motivation or<br />
DDM. Aboulia falls in the middle of the spectrum of<br />
diminished motivation, with apathy being less extreme and<br />
akinetic mutism being more extreme than aboulia. A<br />
patient with aboulia is unable to act or make decisions<br />
independently. d It may range in severity from subtle bl to<br />
overwhelming. It is also known as Blocq's disease (which<br />
also refers to abasia and astasia‐abasia). Abulia was<br />
originally considered to be a disorder of the will. [<br />
29
19/07/54<br />
Aboulia has been known to clinicians since 1838. However, in the time since its<br />
inception, the definition of aboulia has been subjected to many different forms, some<br />
even contradictory with previous ones. Aboulia has been described as a loss of drive,<br />
expression, loss of behavior and speech output, slowing and prolonged speech latency,<br />
and reduction of spontaneous thought content and initiative. <strong>The</strong> clinical features most<br />
commonly associated with aboulia are:<br />
1. Difficulty in initiating and sustaining purposeful movements<br />
2. Lack of spontaneous movement<br />
3. Reduced spontaneous movement<br />
4. Increased response‐time to queries<br />
5. Passivity<br />
6. Reduced emotional responsiveness and spontaneity<br />
7. Reduced social interactions<br />
8. Reduced interest in usual pastimes<br />
Especially in patients with progressive dementia, it may affect feeding. Patients may<br />
continue to chew or hold food in their mouths for hours without swallowing it. <strong>The</strong><br />
behavior may be most evident after these patients have eaten part of their meals<br />
and no longer have strong appetites.<br />
Amotivational syndrome is a psychological condition associated<br />
with diminished inspiration to participate in social situations and activities,<br />
with lapses in apathy caused by an external event, situation, substance (or<br />
lack of), relationship, or other cause.<br />
While some have claimed that chronic use of cannabis causes amotivational<br />
syndrome in some users, empirical studies suggest that there is no such thing<br />
as "amotivational syndrome", per se, but that chronic cannabis intoxication<br />
can lead to apathy and amotivation. From a World Health Organization report:<br />
<strong>The</strong> evidence for an "amotivational syndrome" among adults consists largely<br />
of case histories i and observational reports (e.g. Kl Kolanskyand Moore, 1971;<br />
Millman and Sbriglio, 1986). <strong>The</strong> small number of controlled field and<br />
laboratory studies have not found compelling evidence for such a syndrome<br />
(Dornbush, 1974; Negrete, 1983; Hollister, 1986)... (I)t is doubtful that<br />
cannabis use produces a well defined amotivational syndrome. It may be more<br />
parsimonious to regard the symptoms of impaired motivation as symptoms of<br />
chronic cannabis intoxication rather than inventing a new psychiatric<br />
syndrome.<br />
Apathy (also called impassivity or<br />
perfunctoriness) is a state of<br />
indifference, or the suppression of emotions<br />
such as concern, excitement, motivation and<br />
passion. An apathetic individual has an<br />
absence of interest in or concern about<br />
emotional, social, spiritual, philosophical or<br />
physical life.<br />
<strong>The</strong>y may lack a sense of purpose or<br />
meaning in their life. He or she may also<br />
exhibit hb insensibility bl or sluggishness. <strong>The</strong><br />
opposite of apathy is flow. In positive<br />
psychology, apathy is described as a result of<br />
the individual feeling they have much more<br />
than the level of skill required to confront a<br />
challenge. It may also be a result of<br />
perceiving no challenge at all (e.g. the<br />
challenge is irrelevant to them, or conversely,<br />
they have learned helplessness).<br />
Brodmann area 25 (BA25) is an area in the<br />
cerebral cortex of the brain and delineated based on<br />
its cytoarchitectonic characteristics, also called the<br />
subgenual area, area subgenualis or subgenual<br />
cingulate. It is the 25th "Brodmann area" defined by<br />
Korbinian Brodmann (thus its name). BA25 is located in the<br />
cingulate region as a narrow band in the caudal portion of<br />
the subcallosal area adjacent to the paraterminal gyrus. <strong>The</strong><br />
posterior parolfactory sulcus separates the paraterminal<br />
gyrus from BA25. Rostrally it is bound by the prefrontal area<br />
11 of Brodmann. This region is extremely rich in serotonin<br />
transporters and is considered as a governor for a vast<br />
network involving areas like hypothalamus and brain stem,<br />
which influences changes in appetite and sleep; the<br />
amygdala and insula, which affect the mood and anxiety; the<br />
hippocampus, which plays an important role in memory<br />
formation; and some parts of the frontal cortex responsible<br />
for self‐esteem.<br />
One study has noted that BA25 is metabolically overactive in treatmentresistant<br />
depression and has found that chronic deep brain stimulation in the<br />
white matter adjacent to the area is a successful treatment for some patients.<br />
A different study found that metabolic hyperactivity in this area is associated<br />
with poor therapeutic response of persons with Major Depressive Disorder to<br />
cognitive‐behavioral therapy and venlafaxine<br />
Brain: Brodmann area 25<br />
Brodmann area 33, , also known as<br />
pregenual area 33, is a subdivision<br />
of the cytoarchitecturally defined<br />
cingulate region of cerebral cortex. It<br />
is a narrow band located in the<br />
anterior cingulate gyrus adjacent to<br />
the supracallosal gyrus in the depth<br />
of the callosal sulcus.<br />
Cytoarchitecturally it is bounded by<br />
the ventral anterior cingulate area<br />
24 and the supracallosal gyrus<br />
(Brodmann‐1909).<br />
Brain: Brodmann area 33<br />
30
19/07/54<br />
Impairment of social and moral behavior<br />
related to early damage in human prefrontal<br />
cortex<br />
Steven W. Anderson, Antoine Bechara, Hanna Damasio, Daniel Tranel &<br />
Antonio R. Damasio<br />
Department of Neurology, Division of Behavioral Neurology and Cognitive<br />
Neuroscience, <strong>The</strong> <strong>University</strong> of Iowa College of Medicine, Iowa City, Iowa 52242,<br />
USA<br />
<strong>The</strong> long-term consequences of early prefrontal cortex lesions<br />
occurring before 16 months were investigated in two adults. As is the<br />
case when such damage occurs in adulthood, the two early-onset<br />
patients had severely impaired social behavior despite normal basic<br />
cognitive abilities, and showed insensitivity to future consequences of<br />
decisions, defective autonomic responses to punishment contingencies<br />
and failure to respond to behavioral interventions. Unlike adult-onset<br />
patients, however, the two patients had defective social and moral<br />
reasoning, suggesting that the acquisition of complex social<br />
conventions and moral rules had been impaired. Thus early-onset<br />
prefrontal damage resulted in a syndrome resembling psychopathy.<br />
เด็กสมาธิสั้น ซน พฤติกรรมไม่เหมาะสมกับกาลเทศะ<br />
(Attention Deficit/Hyperactivity: AD/HD)<br />
ระบบประสาทสมองใน<br />
เด็กสมาธิสั้น ซน<br />
พฤติกรรมไม่เหมาะสม<br />
กับกาลเทศะ<br />
มีความผิดปกติของ<br />
สารเคมสอประสาทใน<br />
ีสื<br />
่ ส<br />
สมองโดปามีน<br />
(Dopamine)<br />
Prefrontal lobe syndrome<br />
• Personality changes<br />
• Deficits in strategic planning<br />
• Perseveration<br />
• Release of primitive reflexes<br />
• Abulia = general slowing of the intellectual<br />
faculties i.e. apathetic, slow speech etc.<br />
<strong>The</strong> Moral Brain and<br />
decision making<br />
31
19/07/54<br />
Fig. 2. Brain areas exhibiting differences in activity between conditions shown in<br />
three axial slices of a standard brain (28). Slice location is indicated by Talairach (28)<br />
z coordinate. Data are for the main effect of condition in Experiment 1. Colored areas<br />
reflect the thresholded F scores. Images are reversed left to right to follow radiologic convention.<br />
Fig. 1. Effect of condition on activity in brain areas identified in Experiment 1. R,<br />
right; L, left; B, bilateral. Results for the middle frontal gyrus were not replicated in<br />
Experiment 2. <strong>The</strong> moral‐personal condition was significantly different from the<br />
other two conditions in all other areas in both Experiments 1 and 2. In Experiment<br />
1 the medial frontal and posterior cingulate gyri showed significant differences<br />
between the moral‐impersonal and non‐moral conditions. In Experiment 2 only the posterior cingulate gyrus was<br />
significantly different in this comparison. Brodmann's Areas and Talairach (28) coordinates (x, y, z) for each area are as<br />
follows (left to right in graph): 9/10 (1, 52, 17); 31 (-4, -54, 35); 46 (45, 36, 24); 7/40 (-48, -65, 26); 7/40 (50, -57, 20).<br />
Jorge Moll, Roland Zahn, Frank Krueger and Jordan Grafman are at <strong>The</strong> Cognitive<br />
Neuroscience Section, National Institute<br />
of Neurological Disorders and Stroke, Building 10; Room 5C205; MSC 1440, NIH,<br />
Bethesda, Maryland 20892‐1440, USA.<br />
Ricardo de Oliveira‐Souza is at the Cognitive and Behavioral Neuroscience Unit,<br />
LABS‐D’Or Hospital Network, R. Pinheiro Guimaraes 22, 3rd floor, Rio de Janeiro<br />
22281‐080, Brazil.<br />
Correspondence to J.G. e‐mail: grafmanj@ninds.nih.gov<br />
Cognitive Neuroscience Section, NINDS, NIH:<br />
http://intra.ninds.nih.gov/Lab.asp?Org_ID=83<br />
Cognitive and Behavioural Neuroscience Unit,<br />
LABS‐D’Or Hospital Network:<br />
http://www.rededor.com.br/cbnu/<br />
32
19/07/54<br />
<strong>Frontal</strong> lobe and Executive Function<br />
Disorder<br />
33
19/07/54<br />
<strong>The</strong> <strong>Frontal</strong> Lobes Are Considered<br />
Our Emotional Control Center and<br />
Home to Our Personality<br />
<strong>The</strong>re Is No Other Part of the Brain Where<br />
Lesions Can Cause Such a Wide Variety of<br />
Symptoms.<br />
‐ involved in motor function, problem<br />
solving, spontaneity, memory, language,<br />
initiation, judgement, impulse control,<br />
and social iland sexual behavior. bh ‐ extremely vulnerable to injury due to<br />
their location at the front of the cranium,<br />
proximity to the sphenoid wing and their<br />
large size.<br />
<strong>The</strong>re Are Important Asymmetrical Differences<br />
in the <strong>Frontal</strong> Lobes.<br />
<strong>The</strong> left frontal lobe is involved in controlling<br />
language related movement, whereas the<br />
right frontal lobe plays a role in non‐verbal<br />
abilities.<br />
Some researchers emphasize that this rule is<br />
not absolute and that with many people,<br />
both <strong>lobes</strong> are involved in nearly all<br />
behavior.<br />
Disturbance of motor function is typically<br />
characterized by loss of fine movements<br />
and strength of the arms, hands and fingers.<br />
Complex chains of motor movement also seem<br />
to be controlled by the frontal <strong>lobes</strong>.<br />
Patients with frontal lobe damage exhibit little<br />
spontaneous facial expression, which points<br />
to the role of the frontal <strong>lobes</strong> in facial<br />
expression.<br />
Broca's Aphasia, or difficulty in speaking, has<br />
been associated with frontal damage.<br />
34
19/07/54<br />
An interesting phenomenon of frontal lobe<br />
damage is the insignificant effect it can have<br />
on traditional IQ testing.<br />
<strong>Frontal</strong> lobe damage seems to have an<br />
impact on divergent thinking, or flexibility<br />
and problem solving ability.<br />
<strong>The</strong>re is also evidence showing lingering<br />
interference with attention and memory<br />
even after good recovery from a TBI.<br />
One Characteristics of <strong>Frontal</strong> Lobe Damage<br />
Is Difficulty in Interpreting Feedback From the<br />
Environment<br />
Perseverating on a response, risk taking and<br />
non‐compliance with rules, and impaired<br />
associated learning (using external cues<br />
to help guide behavior) are a few<br />
examples of this type of deficit.<br />
<strong>The</strong> frontal <strong>lobes</strong> are also thought to play a<br />
part in our spatial orientation, including<br />
our bodies orientation in space.<br />
One of the Most Common Effects of <strong>Frontal</strong><br />
Damage Can Be a Dramatic Change in Social<br />
Behavior.<br />
A person's personality can undergo<br />
significant changes after an injury to the<br />
frontal <strong>lobes</strong>, especially when both <strong>lobes</strong><br />
are involved.<br />
<strong>The</strong>re are some differences in the left versus<br />
right frontal <strong>lobes</strong> in this area.<br />
Left frontal damage usually manifests as<br />
pseudodepression and right frontal<br />
damage as pseudopsychopathic.<br />
One of the most common effects of frontal<br />
damage can be a dramatic change in social<br />
behavior.<br />
Sexual behavior can also be effected by<br />
frontal lesions.<br />
Orbital frontal damage can introduce<br />
abnormal sexual behavior, while<br />
dorsolateral lesions may reduce sexual<br />
interest.<br />
Some Common Tests for <strong>Frontal</strong> Lobe<br />
Function Are:<br />
Wisconsin Card Sorting (response<br />
inhibition);<br />
Finger Tapping (motor skills);<br />
Token Test (language skills).<br />
<strong>The</strong> Wisconsin Card Sorting Test<br />
(WCST) is a neuropsychological test of "setshifting",<br />
i.e. the ability to display flexibility in the<br />
face of changing schedules of reinforcement.<br />
<strong>The</strong> WCST was written by David A. Grant and Esta A. Berg.<br />
<strong>The</strong> Professional Manual for the WCST was written by Robert<br />
K. Heaton, Gordon J. Chelune, Jack L. Talley, Gary G. Kay, and<br />
Glenn Curtiss.<br />
Clinically, the test is widely used by neuropsychologists,<br />
clinical psychologists, neurologists and psychiatrists in<br />
patients with acquired brain injury, neurodegenerative<br />
disease, or mental tlillness such as schizophrenia. h i It has been In particular, patients with ihlesions<br />
considered a measure of executive function because of its of the dorsolateral frontal lobe<br />
reported sensitivity to frontal lobe dysfunction. As such, the<br />
make a higher number of<br />
WCST allows the clinician to assess the following "frontal"<br />
lobe functions: strategic planning, organized searching,<br />
perseverative errors than control<br />
utilizing environmental feedback to shift cognitive sets, participants.<br />
directing behavior toward achieving a goal, and modulating A recent factor analysis of the WCST<br />
impulsive responding. <strong>The</strong> test can be administered to those has shown these perseverative errors<br />
6.5 years to 89 years of age.<br />
to be the most useful outcome<br />
Although successful completion of the test relies upon a measure in assessing cases. A more<br />
number of intact cognitive functions including attention, sophisticated description of deficits of<br />
working memory, and visual processing, it is loosely termed a this type is "executive dysfunction".<br />
"frontal lobe" test on the basis that patients with any sort of<br />
frontal lobe lesion generally do poorly at the test.<br />
35
19/07/54<br />
Current News:<br />
‐ Multi-Tasking Pinpointed to <strong>Frontal</strong> Lobes.<br />
‐ Two Brain Areas Involved in Doing Math.<br />
‐ Brain Can Compensate for Damage to Language<br />
Systems After Injury.<br />
‐ Injury to <strong>Frontal</strong> Lobes May Affect Appreciation<br />
of Humor.<br />
Finger Tapping (motor skills)<br />
Executive Function Disorder<br />
<strong>The</strong> frontal <strong>lobes</strong> represent a large area<br />
of the brain and brain centers within<br />
the frontal lobe have numerous<br />
interconnections with other parts of<br />
the brain.<br />
<strong>The</strong>se include connections with emotion<br />
and mood centers as well as cognitive<br />
centers.<br />
Executive Function Disorder<br />
This high degree of complexity results in<br />
a number of syndromes that are<br />
associated with lesions of this area.<br />
<strong>The</strong>re are three general anatomical<br />
divisions of the frontal cortex: the<br />
limbic, the precentral and the<br />
prefrontal cortices.<br />
<strong>The</strong> <strong>Frontal</strong> Lobes<br />
<strong>The</strong> precentral cortex consists of areas<br />
that lie immediately before the<br />
central sulcus.<br />
<strong>The</strong>se consist of the primary and<br />
secondary motor control areas.<br />
<strong>The</strong> <strong>Frontal</strong> Lobes<br />
<strong>The</strong> limbic component consists of<br />
the inferior and medial parts of the<br />
cingulate gyrus and the posterior<br />
parts of the orbital frontal areas.<br />
<strong>The</strong>se areas have inter‐ connections<br />
with the amygdala, hippocampus,<br />
thalamus and other parts of the<br />
limbic system.<br />
36
19/07/54<br />
<strong>The</strong> <strong>Frontal</strong> Lobes<br />
<strong>The</strong> prefrontal cortex is anterior to the<br />
motor control areas and comprises the<br />
greater part of the frontal lobe.<br />
It is subdivided into the dorsolateral,<br />
mesial and orbital areas.<br />
<strong>The</strong>se sections are histologically distinct<br />
and probably represent functional<br />
differences.<br />
<strong>The</strong> <strong>Frontal</strong> Lobes<br />
<strong>The</strong> prefrontal cortex is the area<br />
implicated in the studies of<br />
personality and the behavioral<br />
effects of frontal lobe lesions.<br />
It has extensive interconnections<br />
with virtually every other part of<br />
the cerebrum.<br />
<strong>The</strong> <strong>Frontal</strong> Lobes<br />
<strong>The</strong>se include association areas in<br />
the temporal, occipital and parietal<br />
<strong>lobes</strong>, the limbic system, the dorsal<br />
medial nucleus of the thalamus<br />
and the basal ganglia.<br />
<strong>The</strong> <strong>Frontal</strong> Lobes<br />
Bilateral lesion of the inferior medial<br />
sections of the frontal <strong>lobes</strong> causes<br />
emotional and behavioral changes.<br />
Intellectual impairment results from<br />
lesion of the dorso‐ lateral portion of<br />
the frontal <strong>lobes</strong>.<br />
Cognitive Impairment<br />
Although patients with frontal lobe<br />
lesions may have extensive brain<br />
injury and numerous behavioral<br />
changes they will typically score<br />
within the normal range on IQ tests.<br />
Cognitive Impairment<br />
<strong>The</strong> cognitive impairment associated with<br />
frontal lobe lesions involve cognitive<br />
abilities that are not measured by<br />
conventional IQ tests.<br />
<strong>The</strong>se include impairment of hypothesis<br />
testing and abstract reasoning, memory<br />
disorder, attention deficits and difficulty<br />
in initiation of cognitive activity.<br />
37
19/07/54<br />
Impairment of Abstract Reasoning,<br />
Hypothesis Testing and Modulation of<br />
Cognition<br />
<strong>The</strong> overriding function of the frontal areas<br />
is to modulate and control motor function,<br />
emotion, attention and other cognitive<br />
activity.<br />
Virtually all aspects of frontal lobe impairment<br />
can be considered a specific aspect<br />
of a general deficit of control.<br />
Impairment of Abstract Reasoning, Hypothesis<br />
Testing and Modulation of Cognition<br />
In the domain of reasoning, a deficit of<br />
modulation is manifested as poor abstract<br />
reasoning and failure to maintain goal‐<br />
directed behavior.<br />
Abstract reasoning involves making complex<br />
associations between semantic elements<br />
and identifying super‐ordinate categories,<br />
reasoning by general rules and formulating<br />
hypotheses.<br />
Impairment of Abstract Reasoning,<br />
Hypothesis Testing and Modulation of<br />
Cognition<br />
If abstract reasoning is not present then the<br />
patient has difficulty formulating the<br />
super‐ordinate category that subsumes<br />
individual semantic elements.<br />
For example, the patient may not be able to reason that<br />
cars, trains, airplanes and the like, are members of the<br />
category of modes of transportation.<br />
Impairment of Abstract Reasoning,<br />
Hypothesis Testing and Modulation of<br />
Cognition<br />
Another manifestation of abstract reasoning<br />
involves the development of rules that<br />
guide future behavior.<br />
<strong>The</strong> reasoning person identifies common<br />
features and themes in the experiences of<br />
everyday life and formulates general rules<br />
that govern behavior in these situations.<br />
Impairment of Abstract Reasoning,<br />
Hypothesis Testing and Modulation of<br />
Cognition<br />
Rules may also be taught by others but reasoning<br />
people usually verify and endorse<br />
the validity of rules before using them.<br />
People who have sustained injury to the<br />
frontal <strong>lobes</strong> have difficulty formulating<br />
these rules.<br />
Impairment of Abstract Reasoning,<br />
Hypothesis Testing and Modulation<br />
of Cognition<br />
Indeed, even if a rule is given to the patient<br />
there is still great difficulty in using it to<br />
guide bh behavior.<br />
As a result of this general inability to<br />
formulate and use rules, the patient<br />
cannot conceptualize goal states and use<br />
the goals as objectives to guide thought<br />
and actions.<br />
38
19/07/54<br />
Impairment of Abstract Reasoning,<br />
Hypothesis Testing and Modulation of<br />
Cognition<br />
Actions are motivated by very concrete,<br />
superficial goals, such as immediate<br />
gratification of simple impulses.<br />
Hypothesis formulation and testing also involves<br />
the generalization of experiences in the form of<br />
rules or general principles. This cognitive<br />
function is likewise affected by lesions of the<br />
frontal <strong>lobes</strong>.<br />
Disturbance of Behavior and<br />
Personality<br />
Patients with lesions of the ventro‐<br />
medial portion of the frontal <strong>lobes</strong><br />
have a behavioral syndrome<br />
characterized by lack of originality and<br />
creativity, impairment of attention, and a<br />
tendency to display inappropriate<br />
emotions and behavior.<br />
Disturbance of Behavior and<br />
Personality<br />
<strong>The</strong>y have difficulty initiating behavior;<br />
when they do engage in activity, they<br />
may continue the activity it without<br />
t<br />
stopping.<br />
<strong>The</strong>y may only start activity when<br />
prompted by others.<br />
Disturbance of Behavior and<br />
Personality<br />
Although controversial, emotional<br />
disturbance most often results from<br />
lesion of the orbital frontal areas.<br />
<strong>The</strong>se areas have interconnections<br />
with the amygdala and<br />
hypothalamus.<br />
Disturbance of Behavior and<br />
Personality<br />
Emotional disturbances include laughing<br />
or crying in situations inappropriate to<br />
the emotion.<br />
<strong>The</strong> emotional response also appears<br />
superficial and variable.<br />
<strong>The</strong> patient usually has no awareness<br />
that their emotional response is<br />
incorrect or extreme.<br />
Aphasia<br />
39
19/07/54<br />
Language Impairment<br />
Broca's area resides in the left frontal<br />
lobe and lesion of this area produces a<br />
well‐known impairment of language<br />
referred to as Broca's aphasia.<br />
Patients with dorsolateral frontal lobe<br />
lesions may also have additional<br />
language impairment that is distinct<br />
from Broca's aphasia.<br />
Language Impairment<br />
<strong>The</strong> first of these is a general reduction of<br />
language production although language<br />
utterances that are produced are fluent<br />
and maintain correct syntax.<br />
Patients with frontal lobe lesions have<br />
difficulty initiating speech and<br />
maintaining a complex, spontaneous<br />
conversation.<br />
Language Impairment<br />
This is referred to as a deficit in<br />
"verbal fluency", although language<br />
productions are actually fluent.<br />
Language Impairment<br />
Patients with frontal lesions in secondary<br />
motor control areas may also display<br />
mutism.<br />
In particular, lesions of the cingulum often<br />
result in essential mutism.<br />
Here, the patient comprehends language but<br />
cannot produce any vocalizations, including<br />
language.<br />
Language Impairment<br />
This is viewed as a deficit of the basic<br />
motor control of the oral apparatus to<br />
the point where there is no initiation of<br />
activity.<br />
<strong>The</strong> patient is often indifferent to<br />
communication in general and does<br />
not show the frustration characteristic<br />
of Broca's aphasia.<br />
Language Impairment<br />
A pure agraphia is also associated with<br />
lesions of supplemental motor areas of the<br />
left dorsolateral frontal lobe (Exner's<br />
area).<br />
Exner's area (Broadman area #46<br />
46) is above<br />
Broca's area (Broadman area #44,45) and<br />
anterior to the primary motor control area.<br />
In this disorder, the patient is not aphasic<br />
but has difficulty writing and reading.<br />
40
19/07/54<br />
Impairment of Motor Function<br />
<strong>The</strong> highly controlled, volitional components<br />
of motor control are located in the frontal<br />
<strong>lobes</strong> on the dorso‐ lateral surface anterior<br />
to and including the primary motor control<br />
area (the motor strip).<br />
Other components of the motor control<br />
system are the Basal Ganglia and the<br />
Cerebellum.<br />
<strong>The</strong> cortical system in the frontal <strong>lobes</strong> are<br />
involved in the complex control of skeletal<br />
muscles in the execution of actions.<br />
Impairment of Motor Function<br />
Lesions of the dorsolateral frontal areas<br />
results in a number of motor<br />
impairments.<br />
<strong>The</strong>se include:<br />
‐perseveration,<br />
‐inco‐ordination,<br />
‐motor impersistence, and<br />
‐hypokinesia.<br />
Impairment of Motor Function<br />
‐Patients with left frontal lobe lesions<br />
may also have ideomotor apraxia.<br />
-Lesions within hemisphere cause<br />
worse motor impairment of the<br />
opposite extremities.<br />
Impairment of Motor Function<br />
<strong>The</strong>se deficits again involve the<br />
general modulation and control<br />
functions that are characteristic<br />
of the frontal <strong>lobes</strong>.<br />
Impairment of Motor Function<br />
<strong>The</strong>re is some evidence that control is<br />
lateralized to each frontal area.<br />
<strong>The</strong> left hemisphere is dominant for<br />
motor control and the modulation of<br />
language.<br />
<strong>The</strong> right hemisphere is more involved<br />
in the modulation of actions that are<br />
executed in a spatial context, such as<br />
three‐dimensional block construction.<br />
Impairment of Reflexes<br />
<strong>The</strong> frontal <strong>lobes</strong> play the primary role in the<br />
inhibition of fundamental reflexes that<br />
were presumably inherited as part of<br />
primitive brain structures characteristic of<br />
the primate brain.<br />
<strong>The</strong>se reflexes include the grasp reflex and<br />
the snout and sucking reflexes. <strong>The</strong> grasp<br />
reflex is elicited by stroking the skin of the<br />
palm.<br />
41
19/07/54<br />
Impairment of Reflexes<br />
<strong>The</strong> patient grasps the object and has<br />
difficulty releasing the grasp even when<br />
told to attend to the hand and release<br />
the grasp.<br />
Snout and sucking reflexes are elicited i by<br />
stimulating the lips and space between<br />
the upper lip and the nose. Here, the<br />
patient extends the lips outward for the<br />
snout reflex.<br />
Impairment of Reflexes<br />
<strong>The</strong> patient is compelled to suck on<br />
an object placed in the lips when<br />
the suck reflex is present.<br />
<strong>The</strong> frontal <strong>lobes</strong> usually inhibit these<br />
reflexes during childhood and<br />
adulthood. When the frontal lobe is<br />
lesioned, this inhibition is removed<br />
and the reflexes return.<br />
Impairment of Social Behavior<br />
Since frontal lobe lesions result in a<br />
pervasive defect in planning and<br />
modulation of behavior, these patients<br />
have compelling deficits in maintining<br />
appropriate p social responses.<br />
Social perception and action are very<br />
complex. In addition, people do not<br />
have a wide tolerance for social<br />
behavior; even minor deviations in<br />
social behavior are noticeable.<br />
Impairment of Social Behavior<br />
Patients with frontal lobe lesions have great<br />
difficulty generating appropriate<br />
behavioral options in social situations and<br />
then choosing the best alternative.<br />
<strong>The</strong>y also base their behavior on concrete<br />
simple motivations and cannot formulate<br />
or comprehend more complex or abstract<br />
reasons for acting.<br />
Much social behavior requires a complex<br />
and abstract appreciation of the social<br />
setting.<br />
Confabulation and Reduplication<br />
Syndrome<br />
Patients with severe frontal lobe lesions<br />
tend to fabricate quick, impulsive<br />
answers to questions.<br />
Some responses may be quite fanciful<br />
and imaginative. <strong>The</strong> patient cannot<br />
inhibit a response in order to check its<br />
validity.<br />
Confabulation and Reduplication<br />
Syndrome<br />
This tendency to fabricate an answer<br />
is called confabulation.<br />
It is most common among patients<br />
with basal forebrain lesions and<br />
among patients with additional<br />
impairment of memory ability.<br />
42
19/07/54<br />
Confabulation and Reduplication<br />
Syndrome<br />
Another syndrome that is similar to<br />
confabulation is reduplication.<br />
Here, the patient with a frontal lobe<br />
lesion confabulates that the current<br />
environment, usually the hospital, is<br />
actually another place that is similar<br />
to the current setting but has a<br />
different name and location.<br />
Confabulation and Reduplication<br />
Syndrome<br />
<strong>The</strong> patient may even claim that the current<br />
hospital is a university dormitory or an<br />
apartment building.<br />
<strong>The</strong> confabulated place is always somewhere<br />
else and it is usually familiar to the patient,<br />
such as the hospital in the patient's home<br />
town.<br />
<strong>The</strong> patient will often maintain this<br />
confabulation even when confronted with<br />
salient, contradictory information.<br />
Apraxia<br />
Callosal Disconnection<br />
Syndromes<br />
Executive Function Disorder<br />
43
19/07/54<br />
Blood supply<br />
Branches of the middle cerebral artery provide most of<br />
the arterial blood supply for the primary motor cortex.<br />
<strong>The</strong> medial aspect (leg areas) is supplied by branches of<br />
the anterior cerebral artery.<br />
Neural input from the thalamus<br />
<strong>The</strong> primary motor cortex receive thalamic input from<br />
the Ventral lateral nucleus of the Thalamus.<br />
Pathology<br />
Lesions of the precentral gyrus result in paralysis of the<br />
contralateral side of the body (facial palsy, arm‐/leg<br />
monoparesis, hemiparesis) ‐ see upper motor neuron.<br />
44
19/07/54<br />
Schizophrenia<br />
• Schizophrenia is a psychotic disorder involving<br />
disturbance of thought, emotion and behavior<br />
• <strong>The</strong> lifetime prevalence of schizophrenia is<br />
about 1%<br />
– Onset is usually in late adolescence<br />
– Substance abuse is a co‐morbid condition in 50% of<br />
schizophrenia patients<br />
Ch 11.1<br />
Positive Symptoms of Schizophrenia<br />
Negative Symptoms of Schizophrenia<br />
• Positive symptoms involve excesses or distortions<br />
– Disorganized speech (thought disorder)<br />
– Hallucinations are sensory experiences that occur in the<br />
absence of environmental stimulation<br />
• Hallucinations are commonly auditory<br />
– Delusions are beliefs that are contrary to reality<br />
• Persecutory delusions are common<br />
• Negative schizophrenia symptoms are characterized by<br />
behavioral deficits<br />
– Avolition refers to a lack of energy and an inability to persist<br />
in routine activities<br />
– Alogia refers to a reduction in the amount or content of<br />
speech<br />
– Anhedonia is an inability to experience pleasure<br />
– Asociality refers to a severe impairment in social<br />
relationships<br />
Ch 11.2<br />
Ch 11.3<br />
Etiology of Schizophrenia<br />
• Genetic studies using twin, family and adoption<br />
techniques reveal that a predisposition for<br />
schizophrenia is transmitted genetically<br />
Genetic Studies of Schizophrenia<br />
Relation to<br />
Proband<br />
Spouse 1.00<br />
Grandchildren 2.84<br />
Nieces/nephews 2.65<br />
Children 9.35<br />
Siblings 7.30<br />
DZ twins 12.08<br />
MZ twins 44.30<br />
Percentage<br />
Schizophrenic<br />
Ch 11.5<br />
Ch 11.6<br />
45
19/07/54<br />
Biochemistry of Schizophrenia<br />
• Dopamine theory holds that the positive symptoms of<br />
schizophrenia result from excessive activity of<br />
dopamine in brain<br />
– Anti‐schizophrenia drugs block dopamine receptors<br />
• <strong>The</strong> anti‐schizophrenia h i drugs take several weeks to act clinically, i ll yet<br />
rapidly block dopamine receptors<br />
– Ingestion of amphetamine can induce psychosis;<br />
amphetamine causes the release of dopamine from neurons<br />
Ch 11.7<br />
Dopamine <strong>The</strong>ory of Schizophrenia<br />
Brain Pathology in Schizophrenia<br />
• Brains of schizophrenic patients show<br />
– Reduced volume of temporal and frontal cortex<br />
– Enlarged ventricles (reflecting loss of brain cells)<br />
• For 12 of 15 twins, the schizophrenic h i twin could be<br />
identified by enlarged ventricles<br />
– Reduced metabolic activity within prefrontal cortex<br />
(frontal hypoactivation)<br />
Ch 11.8<br />
Ch 11.9<br />
46
19/07/54<br />
DSM‐IV Schizophrenia Categories<br />
• Disorganized schizophrenia involves<br />
– Disorganized speech and flat affect<br />
– A general disruption of behavior<br />
• Catatonic schizophrenia involves<br />
– Prolonged motor immobility states that alternate with<br />
periods of excitability<br />
• Paranoid schizophrenia involves the presence of<br />
prominent delusions including persecution and<br />
grandiosity<br />
Ch 11.4<br />
Early Forms of Treatment<br />
Insulin Coma <strong>The</strong>rapy<br />
Psychosurgery<br />
Electroconvulsive <strong>The</strong>rapy (ECT)<br />
Smooth Pursuit Eye Tracking<br />
Biological Interventions<br />
Neuroleptics<br />
Haldol & Clozapine<br />
Trial and Error<br />
“Extrapyramidal” Side Effects<br />
Tardive Dyskineasia<br />
Akineasia<br />
<strong>The</strong>rapies for Schizophrenia<br />
• Psychosurgery is the intentional destruction of<br />
brain tissue to alter behavior<br />
– Prefrontal lobotomy was used to treat<br />
schizophrenia<br />
• Drug therapies supplanted psychosurgery<br />
– Use of neuroleptic medications to treat positive<br />
symptoms of schizophrenia<br />
• Chlorpromazine was introduced in the US in 1954<br />
Ch 11.10<br />
Psychological Treatments for<br />
Schizophrenia<br />
• Social‐skills training involves teaching behaviors to<br />
interact successfully with others<br />
• Family therapy aims to reduced expressed emotion<br />
(hostility, overly critical)<br />
• Personal therapy aims to reduce expressed emotion,<br />
uses relaxation techniques and teaches social skills<br />
Ch 11.11`<br />
47