

Ramasamy et al. - 1997 - Yield formation in rice in response to drainage an
Ramasamy et al. - 1997 - Yield formation in rice in response to drainage an
Ramasamy et al. - 1997 - Yield formation in rice in response to drainage an

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
S. Ramsamy <strong>et</strong> <strong>al</strong>. /Field Crops Research 51 (<strong>1997</strong>) 65-82 15<br />
Nod<strong>al</strong> root length<br />
8;<br />
l- ‘-.<br />
6.<br />
E 5- &\<br />
f 4.<br />
Aa -- D”<br />
E 3- - Dn+N<br />
Flower<strong>in</strong>g<br />
/<br />
;2- FL . -~I--- .~~ =:+,<br />
0 20 40 60 SO 100 120<br />
Days after tr<strong>an</strong>spl<strong>an</strong>t<strong>in</strong>g<br />
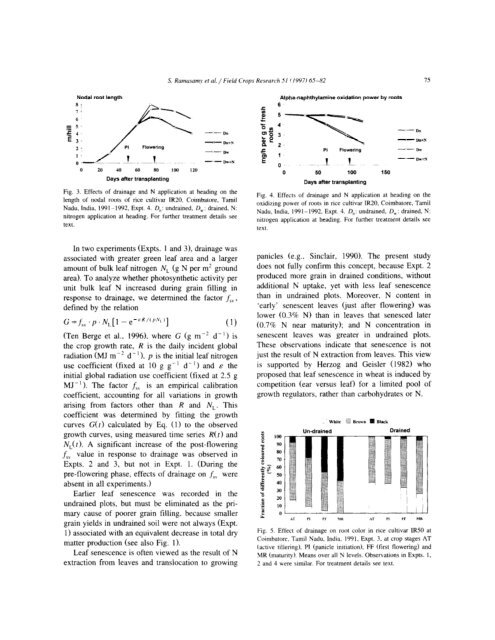
Fig. 3. Effects of dra<strong>in</strong>age <strong>an</strong>d N application at head<strong>in</strong>g on the<br />
length of nod<strong>al</strong> roots of <strong>rice</strong> cultivar IR20, Coimba<strong>to</strong>re, Tamil<br />
Nadu, India, 1991- 1992, Expt. 4. D,: undra<strong>in</strong>ed, D,: dra<strong>in</strong>ed, N:<br />
nitrogen application at head<strong>in</strong>g. For further treatment d<strong>et</strong>ails see<br />
text.<br />
Alpha-naphthylam<strong>in</strong>e oxidation power by roots<br />
0 50 100 150<br />
Days after tr<strong>an</strong>spl<strong>an</strong>t<strong>in</strong>g<br />
Fig. 4. Effects of dra<strong>in</strong>age <strong>an</strong>d N application at head<strong>in</strong>g on the<br />
oxidiz<strong>in</strong>g power of roots <strong>in</strong> <strong>rice</strong> cultivar IR20, Coimba<strong>to</strong>re, Tamil<br />
Nadu, India, 1991-1992. Expt. 4. 0,: undra<strong>in</strong>ed, D,: dra<strong>in</strong>ed, N:<br />
nitrogen application at head<strong>in</strong>g. For further treatment d<strong>et</strong>ails see<br />
text<br />
In two experiments (Expts. 1 <strong>an</strong>d 3), dra<strong>in</strong>age was<br />
associated with greater green leaf area <strong>an</strong>d a larger<br />
amount of bulk leaf nitrogen NL (g N per m’ ground<br />
area). To <strong>an</strong><strong>al</strong>yze wh<strong>et</strong>her pho<strong>to</strong>synth<strong>et</strong>ic activity per<br />
unit bulk leaf N <strong>in</strong>creased dur<strong>in</strong>g gra<strong>in</strong> fill<strong>in</strong>g <strong>in</strong><br />
<strong>response</strong> <strong>to</strong> dra<strong>in</strong>age, we d<strong>et</strong>erm<strong>in</strong>ed the fac<strong>to</strong>r fc,,,<br />
def<strong>in</strong>ed by the relation<br />
G=f,, .p.NL[l -e-EK/(P*v~ ‘1<br />
(Ten Berge <strong>et</strong> <strong>al</strong>., 19961, where G (g m-* d-‘) is<br />
the crop growth rate, R is the daily <strong>in</strong>cident glob<strong>al</strong><br />
radiation (MJ m-’ d-l), p is the <strong>in</strong>iti<strong>al</strong> leaf nitrogen<br />
use coefficient (fixed at 10 g g-’ d-l) <strong>an</strong>d E the<br />
<strong>in</strong>iti<strong>al</strong> glob<strong>al</strong> radiation use coefficient (fixed at 2.5 g<br />
MJ-‘1. The fac<strong>to</strong>r f,, is <strong>an</strong> empiric<strong>al</strong> c<strong>al</strong>ibration<br />
coefficient, account<strong>in</strong>g for <strong>al</strong>l variations <strong>in</strong> growth<br />
aris<strong>in</strong>g from fac<strong>to</strong>rs other th<strong>an</strong> R <strong>an</strong>d N,. This<br />
coefficient was d<strong>et</strong>erm<strong>in</strong>ed by fitt<strong>in</strong>g the growth<br />
curves G(t) c<strong>al</strong>culated by Eq. (1) <strong>to</strong> the observed<br />
growth curves, us<strong>in</strong>g measured time series R(t) <strong>an</strong>d<br />
N,(t). A signific<strong>an</strong>t <strong>in</strong>crease of the post-flower<strong>in</strong>g<br />
f,, v<strong>al</strong>ue <strong>in</strong> <strong>response</strong> <strong>to</strong> dra<strong>in</strong>age was observed <strong>in</strong><br />
Expts. 2 <strong>an</strong>d 3, but not <strong>in</strong> Expt. 1. (Dur<strong>in</strong>g the<br />
pre-flower<strong>in</strong>g phase, effects of dra<strong>in</strong>age on f,, were<br />
absent <strong>in</strong> <strong>al</strong>l experiments.)<br />
Earlier leaf senescence was recorded <strong>in</strong> the<br />
undra<strong>in</strong>ed plots, but must be elim<strong>in</strong>ated as the primary<br />
cause of poorer gra<strong>in</strong> fill<strong>in</strong>g, because sm<strong>al</strong>ler<br />
gra<strong>in</strong> yields <strong>in</strong> undra<strong>in</strong>ed soil were not <strong>al</strong>ways (Expt.<br />
1) associated with <strong>an</strong> equiv<strong>al</strong>ent decrease <strong>in</strong> <strong>to</strong>t<strong>al</strong> dry<br />
matter production (see <strong>al</strong>so Fig. 1).<br />
Leaf senescence is often viewed as the result of N<br />
extraction from leaves <strong>an</strong>d tr<strong>an</strong>slocation <strong>to</strong> grow<strong>in</strong>g<br />
(1)<br />
p<strong>an</strong>icles (e.g., S<strong>in</strong>clair, 1990). The present study<br />
does not fully confirm this concept, because Expt. 2<br />
produced more gra<strong>in</strong> <strong>in</strong> dra<strong>in</strong>ed conditions, without<br />
addition<strong>al</strong> N uptake, y<strong>et</strong> with less leaf senescence<br />
th<strong>an</strong> <strong>in</strong> undra<strong>in</strong>ed plots. Moreover, N content <strong>in</strong><br />
‘early’ senescent leaves (just after flower<strong>in</strong>g) was<br />
lower (0.3% N) th<strong>an</strong> <strong>in</strong> leaves that senesced later<br />
(0.7% N near maturity); <strong>an</strong>d N concentration <strong>in</strong><br />
senescent leaves was greater <strong>in</strong> undra<strong>in</strong>ed plots.<br />
These observations <strong>in</strong>dicate that senescence is not<br />
just the result of N extraction from leaves. This view<br />
is supported by Herzog <strong>an</strong>d Geisler (1982) who<br />
proposed that leaf senescence <strong>in</strong> wheat is <strong>in</strong>duced by<br />
comp<strong>et</strong>ition (ear versus leaf) for a limited pool of<br />
growth regula<strong>to</strong>rs, rather th<strong>an</strong> carbohydrates or N.<br />
Un-dra<strong>in</strong>ed<br />
Dra<strong>in</strong>ed<br />
AT PI T-F MR<br />
Fig. 5. Effect of dra<strong>in</strong>age on root color <strong>in</strong> <strong>rice</strong> cultivar IR50 at<br />
Coimba<strong>to</strong>re, Tamil Nadu. India, 1991. Expt. 3. at crop stages AT<br />
(active tiller<strong>in</strong>g), PI (p<strong>an</strong>icle imtiation). FF (first flower<strong>in</strong>g) <strong>an</strong>d<br />
MR (maturity). Me<strong>an</strong>s over <strong>al</strong>l N levels. Observations <strong>in</strong> Expts. 1,<br />
2 <strong>an</strong>d 4 were similar. For treatment d<strong>et</strong>ails see text.














