118 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 322 men ovale usually present), <strong>and</strong> not in contact with rostral tympanic process <strong>of</strong> petrosal. Anterior limb <strong>of</strong> ectotympanic suspended indirectly from basicranium (by malleus). Stapes usually triangular with large obturator foramen but sometimes columellar <strong>and</strong> imperforate (varies within species). Fenestra cochleae exposed, not concealed by rostral <strong>and</strong> caudal tympanic processes <strong>of</strong> petrosal. Paroccipital process large, erect (not adnate to petrosal) <strong>and</strong> projecting ventrally. Dorsal margin <strong>of</strong> foramen magnum bordered by exoccipitals only, incisura occipitalis absent. Two mental foramina usually present on lateral surface <strong>of</strong> each hemim<strong>and</strong>ible (one foramen is present unilaterally in a single examined specimen <strong>of</strong> D. albiventris); m<strong>and</strong>ibular angular process acute <strong>and</strong> strongly inflected medially. Unworn crowns <strong>of</strong> I2–I5 asymmetrical (‘‘incisiform’’), with much longer anterior than posterior cutting edges. Upper canine (C1) alveolus in premaxillary-maxillary suture; C1 simple, without accessory cusps. First upper premolar (P1) smaller than posterior premolars, but well-formed <strong>and</strong> not vestigial; third upper premolar (P3) taller than second (P2); P3 with posterior cutting edge only; upper milk premolar (dP3) large <strong>and</strong> molariform. Molars highly carnassialized (postmetacristae conspicuously longer than postprotocristae); relative widths M1 , M2 , M3 . M4 or M3 , M4; centrocrista only weakly inflected labially on M1–M3; ect<strong>of</strong>lexus shallow, indistinct, or absent on M1 <strong>and</strong> M2, but consistently deep <strong>and</strong> distinct on M3; anterolabial cingulum <strong>and</strong> preprotocrista discontinuous (anterior cingulum incomplete) on M3; postprotocrista with carnassial notch. Last upper tooth to erupt is M4. Lower incisors (i1–i4) without distinct lingual cusps. Second lower premolar (p2) much taller than p3; lower milk premolar (dp3) with complete (tricuspid) trigonid. Hypoconid labially salient on m3; hypoconulid twinned with entoconid on m1–m3; entoconid much taller than hypoconulid on m1–m3. DISTRIBUTION: Didelphis is a quintessentially eurytopic genus that ranges from southern Canada throughout most <strong>of</strong> North America (except the Rockies, the northern Great Plains, the Great Basin, the arid southwest, north-central Mexico, <strong>and</strong> Baja California; Hall, 1981), all <strong>of</strong> Central America, <strong>and</strong> most <strong>of</strong> South America (to about 40uS latitude in central Argentina; Flores et al., 2007). South American collection records (mapped by Cerqueira <strong>and</strong> Tribe, 2008) represent almost every non-desert tropical <strong>and</strong> subtropical biome on the continent. REMARKS: Despite contradictory or ambiguous <strong>phylogenetic</strong> results from most sequenced loci analyzed separately (table 14), the monophyly <strong>of</strong> Didelphis is strongly supported by parsimony <strong>and</strong> Bayesian analyses <strong>of</strong> morphology (fig. 27), a concatenated five-gene dataset (fig. 33), <strong>and</strong> combined (nonmolecular + molecular) supermatrices (figs. 35, 36). Revisionary studies <strong>of</strong> the genus Didelphis in its modern sense were initiated by Allen (1901, 1902) <strong>and</strong> continued by Krumbiegel (1941), Gardner (1973), Cerqueira (1985), <strong>and</strong> Lemos <strong>and</strong> Cerqueira (2002). Useful summaries <strong>of</strong> morphological characters that distinguish Didelphis species in local faunas are provided by Mondolfi <strong>and</strong> Pérez-Hernández (1984), Catzeflis et al. (1997), Cerqueira <strong>and</strong> Lemos (2000), Flores <strong>and</strong> Abdala (2001), <strong>and</strong> Ventura et al. (2002). Analyses <strong>of</strong> mitochondrial DNA sequence variation within <strong>and</strong> among South American species were reported by Patton et al. (2000) <strong>and</strong> Patton <strong>and</strong> Costa (2003). Lutreolina Thomas, 1910 Figure 47 CONTENTS: crassicaudata Desmarest, 1804 (including crassicaudis Olfers, 1818; bonaria Thomas, 1923; ferruginea Larrañaga, 1923; lutrilla Thomas, 1923; macroura Desmoulins, 1824; paranalis Thomas, 1923; travassosi Mir<strong>and</strong>o-Ribeiro, 1936; <strong>and</strong> turneri Günther, 1879). MORPHOLOGICAL DESCRIPTION: Combined length <strong>of</strong> adult head <strong>and</strong> body ca. 240– 345 mm; adult weight ca. 300–800 g). Rhinarium with one ventrolateral groove on each side <strong>of</strong> median sulcus; head entirely pale <strong>and</strong> unmarked (dark circumocular mask, pale supraocular spots, <strong>and</strong> dark midrostral stripe absent); throat gl<strong>and</strong> absent. Dorsal pelage unpatterned, usually some shade <strong>of</strong> yellowish
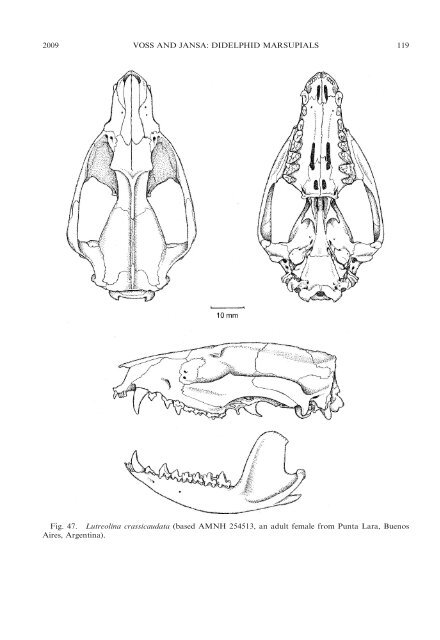
2009 VOSS AND JANSA: DIDELPHID MARSUPIALS 119 Fig. 47. Lutreolina crassicaudata (based AMNH 254513, an adult female from Punta Lara, Buenos Aires, Argentina).