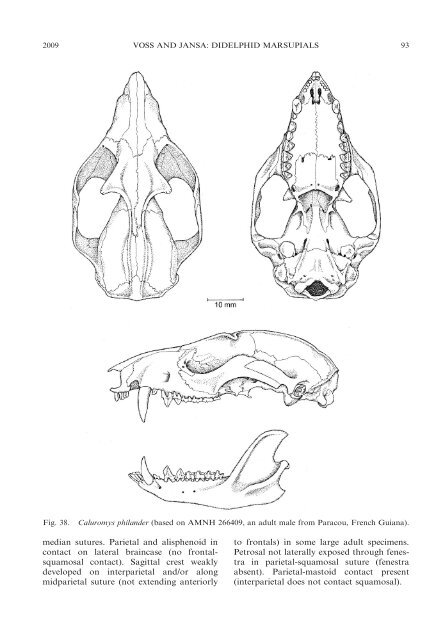
92 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 322 synapomorphies when this gene is eventually sequenced for Caluromysiops. Although most recent authors (e.g., Reig et al., 1985, 1987; Kirsch <strong>and</strong> Palma, 1995; McKenna <strong>and</strong> Bell, 1997; Gardner, 2005, 2008) have included Glironia in the Caluromyinae (or Caluromyidae), that genus appears to represent an ancient lineage <strong>of</strong> basal <strong>didelphid</strong>s that may not be closely related to Caluromys + Caluromysiops (see Jansa <strong>and</strong> Voss, 2000; this report, above). The authorship <strong>of</strong> familygroup names based on Caluromys is <strong>of</strong>ten attributed to Kirsch (1977b), but no such name in that work fulfills the technical criteria for nomenclatural availability (ICZN, 1999: Article 13). Apparently, the first explicit statement <strong>of</strong> characters purported to differentiate this taxon from other opossums appeared in Reig et al. (1987: 72). Caluromys J.A. Allen, 1900 Figure 38 CONTENTS: derbianus Waterhouse, 1841 (including antioquiae Matschie, 1917; aztecus Thomas, 1913; canus Matschie, 1917; centralis Hollister, 1914; fervidus Thomas, 1913; guayanus Thomas, 1899; nauticus Thomas, 1913; pallidus Thomas, 1899; pictus Thomas, 1913; pulcher Matschie, 1917; pyrrhus Thomas, 1901; <strong>and</strong> senex Thomas, 1913); lanatus Olfers, 1818 (including bartletti Matschie, 1917; cahyensis Matschie, 1917; cicur Bangs, 1898; hemiurus Mir<strong>and</strong>a-Ribeiro, 1936; jivaro Thomas, 1913; juninensis Matschie, 1917; lanigera Desmarest, 1820; meridensis Matschie, 1917; modesta Mir<strong>and</strong>a-Ribeiro, 1936; nattereri Matschie, 1917; ochropus Wagner, 1842; ornatus Tschudi, 1845; <strong>and</strong> vitalinus Mir<strong>and</strong>a- Ribeiro, 1936); <strong>and</strong> phil<strong>and</strong>er Linnaeus, 1758 (including affinis Wagner, 1842; cajopolin Müller, 1776; cayopollin Schreber, 1777; cayopollin Kerr, 1792; dichurus Wagner, 1842; flavescens Brongniart, 1792; leucurus Thomas, 1904; trinitatis Thomas, 1894; <strong>and</strong> venezuelae Thomas, 1903). These taxa are currently allocated to two subgenera, Caluromys J.A. Allen, 1900 (containing only C. phil<strong>and</strong>er), <strong>and</strong> Mallodelphys Thomas, 1920 (containing C. derbianus <strong>and</strong> C. lanatus), that can be distinguished unambiguously by the traits noted below. MORPHOLOGICAL DESCRIPTION: Combined length <strong>of</strong> adult head <strong>and</strong> body ca. 210– 300 mm; adult weight ca. 190–500 g. Rhinarium with two ventrolateral grooves on each side <strong>of</strong> median sulcus; dark circumocular mask present; pale supraocular spot absent; dark midrostral stripe present; throat gl<strong>and</strong> absent. Dorsal pelage unpatterned, grayish or reddish brown, or indistinctly mottled with same colors; dorsal underfur grayish; dorsal guard hairs short <strong>and</strong> inconspicuous; ventral fur gray based or self-colored. Manual digit IV longer than other manual digits; manual claws longer than fleshy apical pads <strong>of</strong> digits; dermatoglyph-bearing manual plantar pads present; central palmar epithelium smooth or sparsely tuberculate; carpal tubercles absent. Pedal digits unwebbed; pedal digit IV longer than other pedal digits; plantar surface <strong>of</strong> heel naked. Pouch present, consisting <strong>of</strong> separate lateral skin folds opening medially (subgenus Caluromys) or a deep pocket opening anteriorly (subgenus Mallodelphys); known mammary formulae apparently 2–0–2 5 4 (in C. lanatus) <strong>and</strong> 3–1–3 5 7 (in C. phil<strong>and</strong>er); cloaca present. Tail much longer than combined length <strong>of</strong> head <strong>and</strong> body, slender <strong>and</strong> muscular (not incrassate); body fur extends onto tail base to the same extent dorsally as ventrally (subgenus Caluromys) or much further dorsally than ventrally (subgenus Mallodelphys); naked caudal integument dark (grayish or brownish) basally <strong>and</strong> whitish distally with mottled transition; caudal scales in spiral series, each scale with three subequal bristlelike hairs emerging from distal margin (the central hair <strong>of</strong> each caudal-scale triplet longer than the lateral hairs but not grossly swollen or petiolate); naked ventral caudal surface modified for prehension, with raised tubercles near base (Mallodelphys only) <strong>and</strong> apical pad bearing dermatoglyphs. Premaxillary rostral process present. Nasals long, extending anteriorly above I1 (concealing nasal orifice from dorsal view), <strong>and</strong> conspicuously widened posteriorly near maxillary-frontal suture. Maxillary turbinals elaborately branched. Lacrimal foramina exposed to lateral view at or near anterior orbital margin, usually two on each side. Large, flattened, triangular postorbital processes <strong>of</strong> frontals present. Left <strong>and</strong> right frontals <strong>and</strong> parietals separated by persistent
2009 VOSS AND JANSA: DIDELPHID MARSUPIALS 93 Fig. 38. Caluromys phil<strong>and</strong>er (based on AMNH 266409, an adult male from Paracou, French Guiana). median sutures. Parietal <strong>and</strong> alisphenoid in contact on lateral braincase (no frontalsquamosal contact). Sagittal crest weakly developed on interparietal <strong>and</strong>/or along midparietal suture (not extending anteriorly to frontals) in some large adult specimens. Petrosal not laterally exposed through fenestra in parietal-squamosal suture (fenestra absent). Parietal-mastoid contact present (interparietal does not contact squamosal).