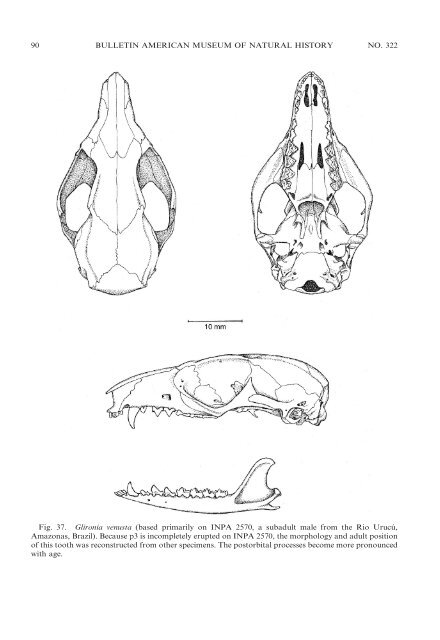
90 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 322 Fig. 37. Glironia venusta (based primarily on INPA 2570, a subadult male from the Rio Urucú, Amazonas, Brazil). Because p3 is incompletely erupted on INPA 2570, the morphology <strong>and</strong> adult position <strong>of</strong> this tooth was reconstructed from other specimens. The postorbital processes become more pronounced with age.
2009 VOSS AND JANSA: DIDELPHID MARSUPIALS 91 INPA 2570) but not others (e.g., FMNH 41440). Upper canine (C1) alveolus in premaxillary-maxillary suture; C1 simple, without accessory cusps. First upper premolar (P1) present, smaller than more posterior premolars but well formed <strong>and</strong> not vestigial; second <strong>and</strong> third upper premolars (P2 <strong>and</strong> P3) subequal in height; P3 with both anterior <strong>and</strong> posterior cutting edges. Molars weakly carnassialized (postmetacristae longer than postprotocristae); relative widths M1 , M2 , M3 . M4; centrocrista weakly inflected labially on M1–M3; ect<strong>of</strong>lexus absent or indistinct on M1 <strong>and</strong> M2, distinct but usually shallow on M3; anterolabial cingulum continuous with preprotocrista (complete anterior cingulum present) on M3; postprotocrista without carnassial notch. Last upper tooth to erupt is P3. Lower incisors (i1–i4) with distinct lingual cusps. Lower canine (c1) erect, acutely pointed, <strong>and</strong> simple (without a posterior accessory cusp). Second lower premolar (p2) taller than p3; morphology <strong>of</strong> lower milk premolar (dp3) unknown (no juvenile specimens examined). Hypoconid labially salient on m3; hypoconulids twinned with entoconids on m1–m3; entoconids much taller than hypoconulids on m1–m3. DISTRIBUTION: Glironia is currently known from fewer than two dozen specimens collected at widely scattered localities between 300 <strong>and</strong> 1000 m above sea level in Brazil (Amazonas, Mato Grosso, Pará, Rodônia), eastern Ecuador, eastern Peru, <strong>and</strong> eastern Bolivia (Díaz <strong>and</strong> Willig, 2004; Santos-Filho et al., 2007; Barkley, 2008). Although the genus probably also occurs in southeastern Colombia, no specimens are known to have been collected there (Díaz <strong>and</strong> Willig, 2004). Most records accompanied by definite habitat information are from primary or secondary Amazonian rainforest, but a single specimen was recently reported from the upper Paraguay Basin (Santos-Filho et al., 2007), <strong>and</strong> several Bolivian records are from dry forest (Tarifa <strong>and</strong> Anderson, 1997; Emmons, 1998). REMARKS: Only the type species <strong>of</strong> Glironia is currently recognized as valid, but it seems probable that additional taxa are represented among the material now preserved in museum collections. Subfamily Caluromyinae Reig et al., 1987 CONTENTS: Caluromys <strong>and</strong> Caluromysiops. DIAGNOSIS: Caluromyines can be distinguished from other confamilial taxa by their long fourth manual digit (dIII is the longest manual digit, or dIII <strong>and</strong> dIV are subequal in all other <strong>didelphid</strong>s); a completely ossified palate (maxillopalatine <strong>and</strong> sometimes additional palatal fenestrae are consistently present in most other <strong>didelphid</strong>s); lack <strong>of</strong> a transverse canal foramen (transverse canal foramina are almost invariably present in all other <strong>didelphid</strong>s); an alisphenoid tympanic process that contacts or closely approximates the rostral tympanic process <strong>of</strong> the petrosal (the alisphenoid tympanic process <strong>and</strong> the rostral tympanic process <strong>of</strong> the petrosal are widely separated in other <strong>didelphid</strong>s); an indirectly suspended anterior limb <strong>of</strong> the ectotympanic (the anterior limb <strong>of</strong> the ectotympanic is directly suspended from the skull in most other <strong>didelphid</strong>s); a fenestra cochleae that is concealed in a sinus formed by the rostral <strong>and</strong> caudal tympanic processes <strong>of</strong> the petrosal (the fenestra cochleae is exposed in most other <strong>didelphid</strong>s); a blunt <strong>and</strong> weakly inflected angular process (the m<strong>and</strong>ibular angle is acute <strong>and</strong> strongly inflected in all other <strong>didelphid</strong>s); an upper canine alveolus that is completely contained by the maxilla (C1 occupies the premaxillarymaxillary suture in all other <strong>didelphid</strong>s); a vestigial or absent first upper premolar (P1 is present <strong>and</strong> nonvestigial in all other <strong>didelphid</strong>s); a tall second upper premolar that much exceeds P3 in height (P2 is either subequal to or smaller than P3 in most other <strong>didelphid</strong>s); <strong>and</strong> lack <strong>of</strong> an ect<strong>of</strong>lexus on all <strong>of</strong> the upper molars (a distinct ect<strong>of</strong>lexus is present, at least on M3, in all other <strong>didelphid</strong>s). REMARKS: Caluromyine monophyly is strongly supported by morphological characters (appendix 5) <strong>and</strong> IRBP gene sequences (fig. 28), <strong>and</strong> this clade is recovered with strong support in analyses <strong>of</strong> concatenated genes (fig. 33) <strong>and</strong> total evidence (nonmolecular + molecular characters; figs. 35, 36) despite the overall incompleteness <strong>of</strong> our data from Caluromysiops (table 2). Two deletions at the DMP1 locus that currently optimize as synapomorphies <strong>of</strong> Caluromys (fig. 29) might prove to be caluromyine