138 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 322 combined datasets that include nonmolecular <strong>and</strong> molecular characters (figs. 35, 36), <strong>and</strong> by Bayesian analyses <strong>of</strong> nonmolecular characters (fig. 27) <strong>and</strong> RAG1 sequences (fig. 30). Spirally arranged caudal scales, the petiolate morphology <strong>of</strong> the central hair in each caudal-scale triplet, the presence <strong>of</strong> a rostral process <strong>of</strong> the premaxillae, <strong>and</strong> unique deletions at the DMP1 <strong>and</strong> BRCA1 loci optimize as unambiguous generic synapomorphies (appendix 5; figs. 29, 31). As noted elsewhere, Kirsch <strong>and</strong> Palma’s (1995) suggestion that Marmosops is paraphyletic was based on misidentified voucher material (Voss <strong>and</strong> Jansa, 2003: 57). Discrepancies between our species-level synonymies for Marmosops <strong>and</strong> those in Gardner (2005) are explained by Voss et al. (2004b). Few <strong>of</strong> the currently recognized species have received critical revisionary attention, <strong>and</strong> it seems likely that several widespread taxa (e.g., M. fuscatus, M. noctivagus, <strong>and</strong> M. impavidus) will prove to be composite. For regional revisions <strong>of</strong> Marmosops see Mustrangi <strong>and</strong> Patton (1997), Patton et al. (2000), <strong>and</strong> Voss et al. (2001, 2004b). Thylamys Gray, 1843 Figure 54 CONTENTS: cinderella Thomas, 1902 (including sponsorius Thomas, 1921); elegans Waterhouse, 1839 (including coquimbensis Tate, 1931; <strong>and</strong> soricinus Philippi, 1894); karimii Petter, 1968; macrurus Olfers, 1818 (including griseus Desmarest, 1827); pallidior Thomas, 1902; pusillus Desmarest, 1804 (including bruchi Thomas, 1921; citellus Thomas, 1912; nanus Olfers, 1818; <strong>and</strong> verax Thomas, 1921); tatei H<strong>and</strong>ley, 1957; velutinus Wagner, 1842 (including pimelurus Reinhardt, 1851); <strong>and</strong> venustus Thomas, 1902 (including janetta Thomas, 1926). MORPHOLOGICAL DESCRIPTION: Combined length <strong>of</strong> adult head <strong>and</strong> body ca. 75– 140 mm; adult weight ca. 10–65 g. Rhinarium with two ventrolateral grooves on each side <strong>of</strong> median sulcus in most examined species (but T. pallidior has only a single ventrolateral groove on each side); dark circumocular mask present; pale supraocular spot absent; dark midrostral stripe absent; throat gl<strong>and</strong> present in fully adult males, <strong>and</strong> apparently also in adult females <strong>of</strong> some species (Carmignotto <strong>and</strong> Monfort, 2006). Dorsal pelage usually grayish <strong>and</strong> distinctly darker middorally than laterally (‘‘tricolored’’), but dorsal fur brownish <strong>and</strong> without conspicuous patterning in T. karimii <strong>and</strong> T. velutinus (Carmignotto <strong>and</strong> Monfort, 2006); dorsal hair bases dark gray; dorsal guard hairs short <strong>and</strong> inconspicuous; ventral fur self-white in some species (e.g., T. pusillus), wholly or partly gray-based whitish or yellowish in others (e.g., T. venustus). Manus mesaxonic (dIII . dIV); manual claws about as long as fleshy apical pads in most species (but claws extend well beyond apical pads in T. karimii <strong>and</strong> T. velutinus; Carmignotto <strong>and</strong> Monfort, 2006); dermatoglyph-bearing manual plantar pads present in most species; central palmar epithelium densely covered with small convex tubercles; carpal tubercles absent in both sexes. Pedal digits unwebbed; dIV longer than other pedal digits; plantar surface <strong>of</strong> heel coarsely furred. Pouch absent; mammae varying among species from 4–1–4 5 9 (in T. karimii; Carmignotto <strong>and</strong> Monfort, 2006) to 7–1–7 5 15 (in T. elegans; Tate, 1933), including ‘‘pectoral’’ teats (when the total number <strong>of</strong> mammae $ 11); cloaca present. Tail longer than combined length <strong>of</strong> head <strong>and</strong> body in most species (except in T. karimii <strong>and</strong> T. velutinus; Carmignotto <strong>and</strong> Monfort, 2006); apparently always incrassate (but quantity <strong>of</strong> stored fat may vary seasonally); furred only at base (to about the same extent dorsally as ventrally); unfurred caudal integument bicolored (dark above, pale below) in most species <strong>and</strong> parti-colored in some (T. macrurus has an all-white tail tip); tail scales in annular series, each scale with three subequal bristlelike hairs emerging from distal margin; ventral caudal surface modified for prehension distally (with naked median groove <strong>and</strong> apical pad bearing dermatoglyphs) in most species, but ventral prehensile surface reduced or absent in T. karimii <strong>and</strong> T. velutinus (Carmignotto <strong>and</strong> Monfort, 2006). Premaxillary rostral process absent. Nasals long, extending anteriorly above or beyond I1 (concealing nasal orifice in dorsal view), <strong>and</strong> uniformly narrow (with subparallel lateral margins) in most specimens. Maxillary turbinals elaborately branched. Two lacrimal
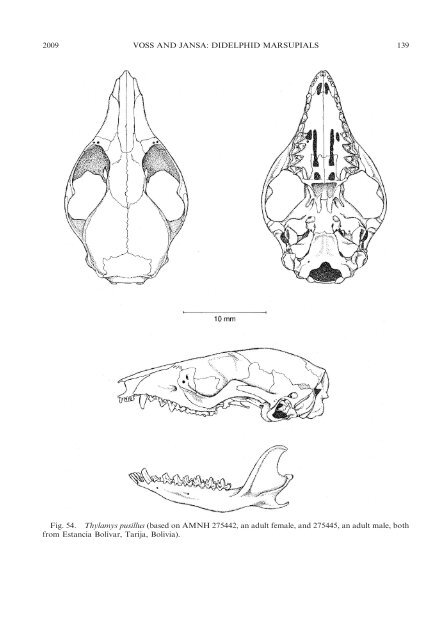
2009 VOSS AND JANSA: DIDELPHID MARSUPIALS 139 Fig. 54. Thylamys pusillus (based on AMNH 275442, an adult female, <strong>and</strong> 275445, an adult male, both from Estancia Bolívar, Tarija, Bolivia).