

Mekanismer for brun- og hvidmuld svampes nedbrydning af tr - Nature
Mekanismer for brun- og hvidmuld svampes nedbrydning af tr - Nature
Mekanismer for brun- og hvidmuld svampes nedbrydning af tr - Nature

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
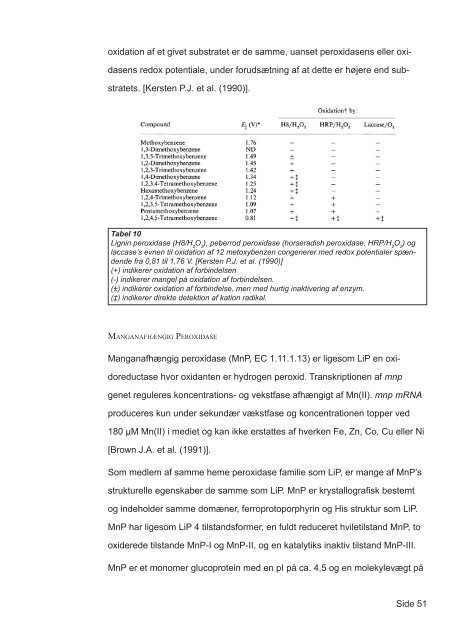
oxidation <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> et givet subs<strong>tr</strong>atet er de samme, uanset peroxidasens eller oxi-<br />
dasens redox potentiale, under <s<strong>tr</strong>ong>for</s<strong>tr</strong>ong>udsætning <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> at dette er højere end sub-<br />
s<strong>tr</strong>atets. [Kersten P.J. et al. (1990)].<br />
Tabel 10<br />
Lignin peroxidase (H8/H 2 O 2 ), peberrod peroxidase (horseradish peroxidase, HRP/H 2 O 2 ) <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong><br />
laccase’s evnen til oxidation <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> 12 metoxybenzen congenerer med redox potentialer spændende<br />
fra 0,81 til 1,76 V. [Kersten P.J. et al. (1990)]<br />
(+) indikerer oxidation <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> <s<strong>tr</strong>ong>for</s<strong>tr</strong>ong>bindelsen<br />
(-) indikerer mangel på oxidation <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> <s<strong>tr</strong>ong>for</s<strong>tr</strong>ong>bindelsen.<br />
(±) indikerer oxidation <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> <s<strong>tr</strong>ong>for</s<strong>tr</strong>ong>bindelse, men med hurtig inaktivering <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> enzym.<br />
(‡) indikerer direkte detektion <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> kation radikal.<br />
Ma n g a n a f h æ n g i g PE r o x i d a s E<br />
Mangan<s<strong>tr</strong>ong>af</s<strong>tr</strong>ong>hængig peroxidase (MnP, EC 1.11.1.13) er ligesom LiP en oxi-<br />
doreductase hvor oxidanten er hydr<s<strong>tr</strong>ong>og</s<strong>tr</strong>ong>en peroxid. Transkriptionen <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> mnp<br />
genet reguleres koncen<strong>tr</strong>ations- <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> vekstfase <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong>hængigt <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> Mn(II). mnp mRNA<br />
produceres kun under sekundær vækstfase <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> koncen<strong>tr</strong>ationen topper ved<br />
180 μM Mn(II) i mediet <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> kan ikke erstattes <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> hverken Fe, Zn, Co, Cu eller Ni<br />
[Brown J.A. et al. (1991)].<br />
Som medlem <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> samme heme peroxidase familie som LiP, er mange <s<strong>tr</strong>ong>af</s<strong>tr</strong>ong> MnP’s<br />
s<strong>tr</strong>ukturelle egenskaber de samme som LiP. MnP er krystall<s<strong>tr</strong>ong>og</s<strong>tr</strong>ong>r<s<strong>tr</strong>ong>af</s<strong>tr</strong>ong>isk bestemt<br />
<s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> indeholder samme domæner, ferroprotoporphyrin <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> His s<strong>tr</strong>uktur som LiP.<br />
MnP har ligesom LiP 4 tilstands<s<strong>tr</strong>ong>for</s<strong>tr</strong>ong>mer, en fuldt reduceret hviletilstand MnP, to<br />
oxiderede tilstande MnP-I <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> MnP-II, <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> en katalytiks inaktiv tilstand MnP-III.<br />
MnP er et monomer glucoprotein med en pI på ca. 4,5 <s<strong>tr</strong>ong>og</s<strong>tr</strong>ong> en molekylevægt på<br />
Side 51