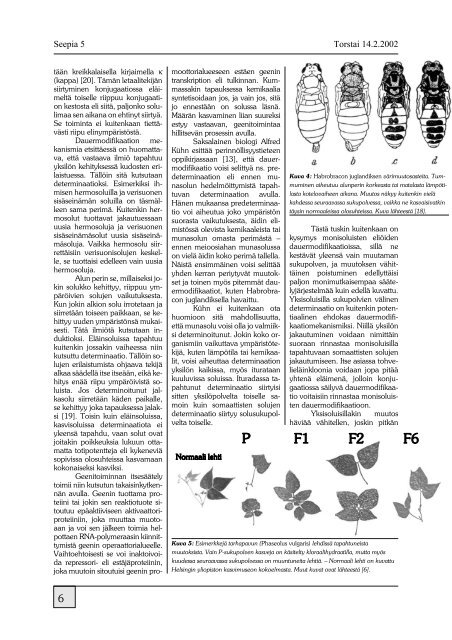
<strong>Seepia</strong> 5Torstai 14.2.2002tään kreikkalaisella kirjaimella κ(kappa) [20]. Tämän letaalitekijänsiirtyminen konjugaatiossa eläimeltätoiselle riippuu konjugaationkestosta eli siitä, paljonko solulimaasen aikana on ehtinyt siirtyä.Se toiminta ei kuitenkaan tiettävästiriipu elinympäristöstä.Dauermodifikaation mekanismiaetsittäessä on huomattava,että vastaava ilmiö tapahtuuyksilön kehityksessä kudosten erilaistuessa.Tällöin sitä kutsutaandeterminaatioksi. Esimerkiksi ihmisenhermosoluilla ja verisuonensisäseinämän soluilla on täsmälleensama perimä. Kuitenkin hermosoluttuottavat jakautuessaanuusia hermosoluja ja verisuonensisäseinämäsolut uusia sisäseinämäsoluja.Vaikka hermosolu siirrettäisiinverisuonisolujen keskelle,se tuottaisi edelleen vain uusiahermosoluja.Alun perin se, millaiseksi jokinsolukko kehittyy, riippuu ympäröiviensolujen vaikutuksesta.Kun jokin alkion solu irrotetaan jasiirretään toiseen paikkaan, se kehittyyuuden ympäristönsä mukaisesti.Tätä ilmiötä kutsutaan induktioksi.Eläinsoluissa tapahtuukuitenkin jossakin vaiheessa niinkutsuttu determinaatio. Tällöin solujenerilaistumista ohjaava tekijäalkaa säädellä itse itseään, eikä kehitysenää riipu ympäröivistä soluista.Jos determinoitunut jalkasolusiirretään käden paikalle,se kehittyy joka tapauksessa jalaksi[19]. Toisin kuin eläinsoluissa,kasvisoluissa determinaatiota eiyleensä tapahdu, vaan solut ovatjoitakin poikkeuksia lukuun ottamattatotipotentteja eli kykeneviäsopivissa olosuhteissa kasvamaankokonaiseksi kasviksi.Geenitoiminnan itsesäätelytoimii niin kutsutun takaisinkytkennänavulla. Geenin tuottama proteiinitai jokin sen reaktiotuote sitoutuuepäaktiiviseen aktivaattoriproteiiniin,joka muuttaa muotoaanja voi sen jälkeen toimia helpottaenRNA-polymeraasin kiinnittymistägeenin operaattorialueelle.Vaihtoehtoisesti se voi inaktoivoidarepressori- eli estäjäproteiinin,joka muutoin sitoutuisi geenin promoottorialueeseenestäen geenintranskription eli tulkinnan. Kummassakintapauksessa kemikaaliasyntetisoidaan jos, ja vain jos, sitäjo ennestään on solussa läsnä.Määrän kasvaminen liian suureksiestyy vastaavan, geenitoimintaahillitsevän prosessin avulla.Saksalainen biologi AlfredKühn esittää perinnöllisyystieteenoppikirjassaan [13], että dauermodifikaatiovoisi selittyä ns. predeterminaationeli ennen munasolunhedelmöittymistä tapahtuvandeterminaation avulla.Hänen mukaansa predeterminaatiovoi aiheutua joko ympäristönsuorasta vaikutuksesta, äidin elimistössäolevista kemikaaleista taimunasolun omasta perimästä –ennen meioosiahan munasolussaon vielä äidin koko perimä tallella.Näistä ensimmäinen voisi selittääyhden kerran periytyvät muutoksetja toinen myös pitemmät dauermodifikaatiot,kuten Habrobraconjuglandiksella havaittu.Kühn ei kuitenkaan otahuomioon sitä mahdollisuutta,että munasolu voisi olla jo valmiiksideterminoitunut. Jokin koko organismiinvaikuttava ympäristötekijä,kuten lämpötila tai kemikaalit,voisi aiheuttaa determinaationyksilön kaikissa, myös iturataankuuluvissa soluissa. Ituradassa tapahtunutdeterminaatio siirtyisisitten yksilöpolvelta toiselle samoinkuin somaattisten solujendeterminaatio siirtyy solusukupolveltatoiselle.Kuva 4: Habrobracon juglandiksen värimuutosasteita. Tummuminenaiheutuu alunperin korkeasta tai matalasta lämpötilastakotelovaiheen aikana. Muutos näkyy kuitenkin vieläkahdessa seuraavassa sukupolvessa, vaikka ne kasvaisivatkintäysin normaaleissa olosuhteissa. Kuva lähteestä [18].Tästä tuskin kuitenkaan onkysymys monisoluisten eliöidendauermodifikaatioissa, sillä nekestävät yleensä vain muutamansukupolven, ja muutoksen vähittäinenpoistuminen edellyttäisipaljon monimutkaisempaa säätelyjärjestelmääkuin edellä kuvattu.Yksisoluisilla sukupolvien välinendeterminaatio on kuitenkin potentiaalinenehdokas dauermodifikaatiomekanismiksi.Niillä yksilönjakautuminen voidaan nimittäinsuoraan rinnastaa monisoluisillatapahtuvaan somaattisten solujenjakautumiseen. Itse asiassa tohvelieläinklooniavoidaan jopa pitääyhtenä eläimenä, jolloin konjugaatiossasäilyvä dauermodifikaatiovoitaisiin rinnastaa monisoluistendauermodifikaatioon.Yksisoluisillakin muutoshäviää vähitellen, joskin pitkänKuva 5: Esimerkkejä tarhapavun (Phaseolus vulgaris) lehdissä tapahtuneistamuutoksista. Vain P-sukupolven kasveja on käsitelty kloraalihydraatilla, mutta myöskuudessa seuraavassa sukupolvessa on muuntuneita lehtiä. – Normaali lehti on kuvattuHelsingin yliopiston kasvimuseon kokoelmasta. Muut kuvat ovat lähteestä [6].6
<strong>Seepia</strong> 5 Torstai 14.2.2002ajan kuluessa. Vaikka yleensä uskotaankin,ettei dauermodifikaationhäviäminen johdu normaalinympäristön kumoavasta vaikutuksesta,olisi mielestäni varsin mahdollista,että esimerkiksi tohvelieläimilleolisi kehittynyt kyky sopeutuamolempiin suuntiin.Solulima jaevoluutioVaikka tietyt elämän aikanahankitut ominaisuudet voivatkinsiirtyä jälkipolville, dauermodifikaatiossaei kuitenkaan ole kysymyshankittujen ominaisuuksienperiytymisestä siinä mielessä kuinLamarck sen ajatteli. Dauermodifikaatioteivät voi osallistua lajinkehitykseen, sillä ne häviävät ainaennemmin tai myöhemmin.Lisäksi dauermodifikaatio ilmeneevain tietyissä ominaisuuksissa –sellaisissa, joiden säätelyjärjestelmäänse on erikseen evolvoitunut.Monissa tapauksissa dauermodifikaatioon selvästikin luonnon valinnanavulla saavutettu sopeutumajatkuvasti muuttuvaan elinympäristöön.Esimerkiksi tohvelieläintenkyky sopeutua erilaisiinlämpötiloihin ilman geeniperimänmuutosta varmasti parantaa lajinelinkelpoisuutta. Samoin vesikirpulleon hyötyä siitä, että se jo elämänsäalussa tietää mahdollisestavaarasta ja voi aloittaa valmistautumisenheti. Tarhapavun dauermodifikaatiotaon kuitenkin vaikeampaaselittää. Kuinka kasvuhäiriöidenperiytyminen voisi ollalajille edullista?Solulimaperiytyminen kuitenkinmitä ilmeisimmin osallistuuevoluutioon, vaikka sen osuus eiliene läheskään yhtä merkittäväkuin geenien. Solulima sisältää paljongeenien toiminnalle välttämätöntäinformaatiota. Muutoksetepigeneettisessä eli geenien transkriptiostaeli tulkinnasta vastaavassasysteemissä luonnollisestivaikuttavat geenien toimintaan,eikä edes standardina pidetty geneettinenkoodi eli DNA:n emästenja aminohappojen välinen vastaavuusole kaikilla eliöillä sama. Joidenkintutkijoiden mukaan epigeneettisessäsysteemissä tapahtuvillamuutoksilla on paljon luultuasuurempi osuus evoluutiossa [5].Soluliman merkitystä lajien välisissäeroavaisuuksissa on vaikea testata,sillä eri lajien yksilöt eivätyleensä risteydy keskenään. Onkuitenkin selvää, että vaikka tohvelieläimeensiirrettäisiin ihmisen perimä,se ei kehittyisi ihmiseksi.Vaikka sukupolvien välinendeterminaatio ei selvästikään aiheutamonisoluisten eliöiden dauermodifikaatiota,se saattaa kuitenkinselittää varsin suuren osan solulimaperiytymisestä.Jos otetaanhuomioon, kuinka yleinen ilmiödeterminaatio on kudosten erilaistumisessa,voisi kuvitella, että kaikillaeliöillä on lukuisia ominaisuuksia,jotka ovat säilyneet soluissa”determinoituneina” – geneettisentakaisinkytkennän avulla – jomiljoonien vuosien ajan ja säilyvätedelleen niin kauan kuin se on lajilleedullista. Nämäkin ominaisuudetsamoin kuin dauermodifikaatiokykysekä muutokset epigeneettisessäsysteemissä syntyvät mitä ilmeisimminluonnonvalinnan eivätkäsiis lamarckismin mukaisestiympäristön suuntaavan vaikutuksentuloksena.Jostakin syystä modernitkehitysbiologian ja perinnöllisyystieteenoppikirjat ja hakuteokset eivätmainitse dauermodifikaatiotalainkaan. Tiedossani ei ole yhtäänmainintaa varsinaisesta dauermodifikaatiostatieteellisessä kirjallisuudessavuoden 1971 jälkeen[13, 14]. Yhden kerran periytyvistämuutoksista on tosin myöhemminkintehty lukuisia tutkimuksia maternaaliefektinnimellä [15].Referoimani tutkimuksetovat kuitenkin mielestäni olleetperusteellisia, eikä ilmiön olemassaoloaole syytä epäillä. En olemyöskään löytänyt yhtään vakavastiotettavaa kritiikkiä. Vaikkadauermodifikaatio onkin mitä ilmeisimmintodellinen ilmiö, siinäei kuitenkaan missään tapauksessaole kysymys lamarckismista,kuten eräät kirjoittajat (esim. [4])tuntuvat luulevan. Dauermodifikaatioei myöskään ole millään tavoinristiriidassa evoluutioteoriankanssa, sillä muutokset ovat ainageenitoiminnan säätelemiä. Dauermodifikaatiostaei kuitenkaantiedetä paljoa. On sääli, ettei siitäole tehty enempää tutkimusta.Sampo TiensuuKirjallisuutta[1] Agrawal, Anurag A.; Laforsch, Christian& Tollrian, Ralph; Transgenerationalinduction of defences in animals andplants. Nature 401 (1999): 60-63.[2] Caspari, Ernst; Cytoplasmaticinheritance. Advances in Genetics 2(1948).[3] Caspari, Ernst; The role of genes andcytoplasmic particles in differentiation.Annals of the New York Academy ofSciences 60 (1955).[4] Custance, Arthur C.; Science and Faith.The Doorway Papers Series 1988.http://www.custance.org/Library/Volume8/index.html[5] Ho, M.W. & Saunders, P.T.; Beyondneo-darwinism – An epigenetic approach toevolution. Journal of Theoretical Biology,78 (1979): 573-591.[6] Hofmann, Frederick Wenzl; Someattempts to modify the germ plasm ofPhaseolus vulgaris. Genetics 12 (1927):284-294.[7] Jollos, Victor; ExperimentelleUntersuchungen an Infusorien. BiologischZentralblatt. 33 (1913): 222-236.[8] Jollos, Victor; ExperimentelleProtistenstudien – I. Untersuchungen überVariabilität und Vererbung bei Infusorien.Archiv für Protistenkunde 43 (1921):1-222.[9] Jollos, Victor; Dauermodifikationen undMutationen bei Protozoen. Archiv fürProtistenkunde 83 (1934): 197-219.[10] Jollos, Victor; Inherited changes producedby heat-treatment in Drosophilamelanogaster. Genetica 16 (1934):476-494.[11] Kaestner, Hans; Die Wirkung vonTemperaturreizen auf die Pigmentierungund Ihre Nachwirkung in den folgendenGenerationen bei Habrobracon JjuglandisAsh. Wilhelm Roux' Archiv fürEntwicklungsmechanik der Organismen124 (1931).[12] King, Karl; Cytoplasm, Inheritance andMutations. 2001.[13] Kühn, Alfred; Grundriß derVererbungslehre. Quelle & Meyer,Heidelberg 1971.[14] Kühn, Alfred; Lectures in DevelopmentalPhysiology. Springer-Verlag, 1971.[15] Rossiter, MaryCarol; Incidence andconsequences of inherited environmentaleffects. Annual Review of Ecology andSystematics 27 (1996): 451-476.[16] Schuckmann, W. von & Piekarski, G.:Beiträge zum Problem derDauermodifikation bei Protozoen. Archivfür Protistenkunde 93 (1940): 355-416.[17] Sirks, M. J.; Plasmatic inheritance. TheBotanical Review IV/3 (1938): 113-131[18] Schlottke, E.; Über die Variabilität derschwarzen Pigmentierung undihreBeeinflußbarkeit durch Temperaturenbei Habrobracon juglandis. Zeitschrift fürVergleichende Physiologie 3 (1926).[19] Walbot, Virginia; Holder, Nigel;Developmental Biology. Random House,1987.[20] Wichterman, Ralph; The Biology ofParamecium. New York, 1953.7