

Linking Specialisation and Stability of Plant ... - OPUS Würzburg
Linking Specialisation and Stability of Plant ... - OPUS Würzburg
Linking Specialisation and Stability of Plant ... - OPUS Würzburg

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
2.6 appendix 35<br />
Here, S <strong>and</strong> T denote the maximum number <strong>of</strong> seeds or pollinator<br />
<strong>of</strong>fspring, respectively, that a single individual can produce<br />
within one time step. h P <strong>and</strong> h A are half-saturation constants.<br />
All other parameters are as defined in the main text.<br />
Community <strong>Stability</strong><br />
0.001<br />
0.000<br />
−0.001<br />
−0.002<br />
γ ik = 0.99<br />
B=1, Holl I<br />
B=16, Holl I<br />
B=1, Holl II<br />
B=16, Holl II<br />
Community <strong>Stability</strong><br />
1e−03<br />
5e−04<br />
0e+00<br />
γ ik = 0.5<br />
−0.003<br />
−5e−04<br />
0.0 0.2 0.4 0.6 0.8 1.0<br />
Pollinator Specialization<br />
0.0 0.2 0.4 0.6 0.8 1.0<br />
Pollinator Specialization<br />
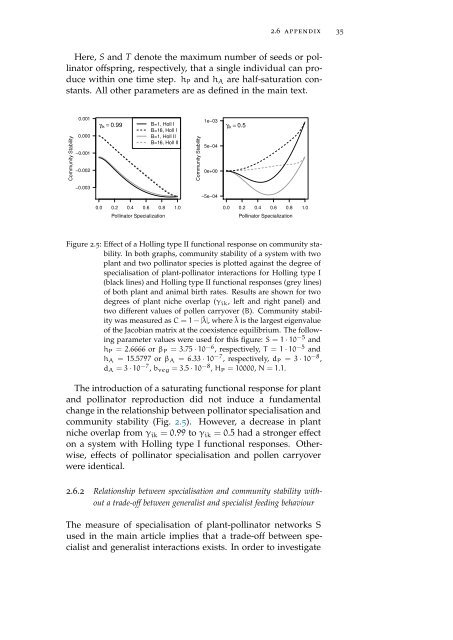
Figure 2.5: Effect <strong>of</strong> a Holling type II functional response on community stability.<br />
In both graphs, community stability <strong>of</strong> a system with two<br />
plant <strong>and</strong> two pollinator species is plotted against the degree <strong>of</strong><br />
specialisation <strong>of</strong> plant-pollinator interactions for Holling type I<br />
(black lines) <strong>and</strong> Holling type II functional responses (grey lines)<br />
<strong>of</strong> both plant <strong>and</strong> animal birth rates. Results are shown for two<br />
degrees <strong>of</strong> plant niche overlap (γ ik , left <strong>and</strong> right panel) <strong>and</strong><br />
two different values <strong>of</strong> pollen carryover (B). Community stability<br />
was measured as C = 1 − |ˆλ|, where ˆλ is the largest eigenvalue<br />
<strong>of</strong> the Jacobian matrix at the coexistence equilibrium. The following<br />
parameter values were used for this figure: S = 1 · 10 −5 <strong>and</strong><br />
h P = 2.6666 or β P = 3.75 · 10 −6 , respectively, T = 1 · 10 −5 <strong>and</strong><br />
h A = 15.5797 or β A = 6.33 · 10 −7 , respectively, d P = 3 · 10 −8 ,<br />
d A = 3 · 10 −7 , b veg = 3.5 · 10 −8 , H P = 10000, N = 1.1.<br />
The introduction <strong>of</strong> a saturating functional response for plant<br />
<strong>and</strong> pollinator reproduction did not induce a fundamental<br />
change in the relationship between pollinator specialisation <strong>and</strong><br />
community stability (Fig. 2.5). However, a decrease in plant<br />
niche overlap from γ ik = 0.99 to γ ik = 0.5 had a stronger effect<br />
on a system with Holling type I functional responses. Otherwise,<br />
effects <strong>of</strong> pollinator specialisation <strong>and</strong> pollen carryover<br />
were identical.<br />
2.6.2 Relationship between specialisation <strong>and</strong> community stability without<br />
a trade-<strong>of</strong>f between generalist <strong>and</strong> specialist feeding behaviour<br />
The measure <strong>of</strong> specialisation <strong>of</strong> plant-pollinator networks S<br />
used in the main article implies that a trade-<strong>of</strong>f between specialist<br />
<strong>and</strong> generalist interactions exists. In order to investigate













