

Anhang i. Simulationsmodell - FG Mikroelektronik, TU Berlin
Anhang i. Simulationsmodell - FG Mikroelektronik, TU Berlin
Anhang i. Simulationsmodell - FG Mikroelektronik, TU Berlin

Sie wollen auch ein ePaper? Erhöhen Sie die Reichweite Ihrer Titel.
YUMPU macht aus Druck-PDFs automatisch weboptimierte ePaper, die Google liebt.
<strong>Anhang</strong><br />
i. <strong>Simulationsmodell</strong><br />
Vm<br />
<strong>Anhang</strong> 107<br />
<strong>Simulationsmodell</strong><br />
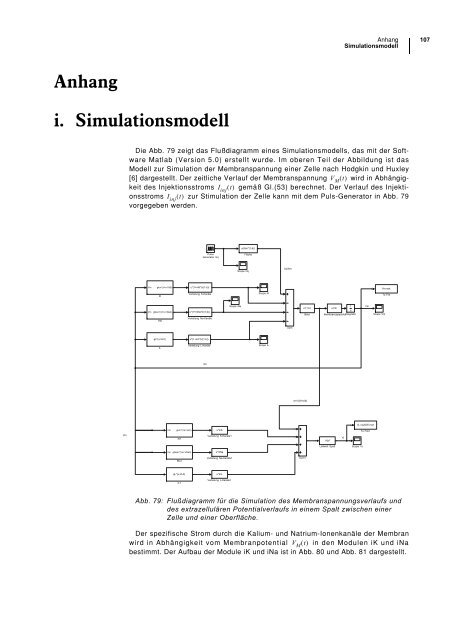
Die Abb. 79 zeigt das Flußdiagramm eines <strong>Simulationsmodell</strong>s, das mit der Software<br />
Matlab (Version 5.0) erstellt wurde. Im oberen Teil der Abbildung ist das<br />
Modell zur Simulation der Membranspannung einer Zelle nach Hodgkin und Huxley<br />
[6] dargestellt. Der zeitliche Verlauf der Membranspannung VM() t wird in Abhängigkeit<br />
des Injektionsstroms Iinj() t gemäß Gl.(53) berechnet. Der Verlauf des Injektionsstroms<br />
Iinj() t zur Stimulation der Zelle kann mit dem Puls-Generator in Abb. 79<br />
vorgegeben werden.<br />
Vm gkm*(Vm-Vk0)<br />
iK<br />
Vm gNam*(Vm-VNa0)<br />
iNa<br />
gL*(u-VL0)<br />
iL<br />
Vm gkm*(Vm-Vk0)<br />
iK1<br />
Vm gNam*(Vm-VNa0)<br />
iNa1<br />
gL*(u-VL0)<br />
iL1<br />
u*(1+mK*b/(1-b))<br />
u*(1+mNa*b/(1-b))<br />
Verteilung Na-Kanäle<br />
u*(1+mL*b/(1-b))<br />
Verteilung L-Kanäle<br />
Pulse<br />
Generator Iinj<br />
Verteilung K-Kanäle<br />
Vm<br />
u*mK<br />
Verteilung K-Kanäle1<br />
u*mNa<br />
Verteilung Na-Kanäle1<br />
u*mL<br />
Verteilung L-Kanäle1<br />
Scope iNa<br />
u/(Am*(1-b))<br />
Fläche<br />
Scope Iinj<br />
Scope iK<br />
Scope iL<br />
Abb. 79: Flußdiagramm für die Simulation des Membranspannungsverlaufs und<br />
des extrazellulären Potentialverlaufs in einem Spalt zwischen einer<br />
Zelle und einer Oberfläche.<br />
Der spezifische Strom durch die Kalium- und Natrium-Ionenkanäle der Membran<br />
wird in Abhängigkeit vom Membranpotential VM() t in den Modulen iK und iNa<br />
bestimmt. Der Aufbau der Module iK und iNa ist in Abb. 80 und Abb. 81 dargestellt.<br />
Iinj/Am<br />
Sum<br />
cm*(dVm/dt)<br />
Sum1<br />
u*(1-b)<br />
Beta<br />
1<br />
u/cm<br />
s<br />
Membrancapazit‰t Integrator<br />
u/gJ<br />
Leitwert Spalt<br />
Vj<br />
Vm<br />
Vj_na2k05.mat<br />
Scope Vj<br />
To File1<br />
Scope Vm<br />
Vm.mat<br />
To File
1<br />
Vm<br />
Vm n<br />
n(Vm)<br />
<strong>Anhang</strong> 108<br />
<strong>Simulationsmodell</strong><br />
Abb. 80: Modul iK. Berechnung des flächenspezifischen Stroms durch die<br />
Kalium-Ionenkanäle in Abhängigkeit der Membranspannung VM() t .<br />
K<br />
K<br />
gK = g0ist die maximale Leitfähigkeit der Kaliumkanäle und VK0 = V0 ist das Nernst-Potential der Kalium-Ionen.<br />
1<br />
Vm<br />
Vm<br />
m(Vm)<br />
Vm h<br />
h(Vm)<br />
m<br />
h<br />
gNa*u^3<br />
gNa*m^3<br />
Abb. 81: Modul iNa. Berechnung des flächenspezifischen Stroms durch die<br />
Natrium-Ionenkanäle in Abhängigkeit der Membranspannung VM() t .<br />
Na<br />
gNa = g0ist die maximale Leitfähigkeit der Natriumkanäle und<br />
Na<br />
VNa0 = V0 ist das Nernst-Potential der Natrium-Ionen.<br />
Die Abhängigkeit der spezifischen Leitfähigkeit von der Membranspannung wurde<br />
anhand der Ratengleichungen von Hudgkin und Huxley Gl.(15) und Gl.(16) berechnet.<br />
Die Faktoren n, m und h ergeben sich durch Integration der Gl.(18) bis Gl.(20),<br />
die in den Abb. 82 bis Abb. 84 grafisch dargestellt sind. Der lineare Anteil des<br />
Membranstroms wird in dem Modul iL berechnet.<br />
Die Verteilung der Natrium, Kalium und ohmschen Kanäle (L-Kanäle) zwischen<br />
Na<br />
K<br />
Kontaktbereich und freier Membran geht durch die Faktoren mNa = μJ , mK = μJ L<br />
und mL = μJ in das Modell ein. Das Verhältnis zwischen der Membranfläche im<br />
Kontaktbereich und freier Membranfläche wird durch den Faktor b = β angegeben.<br />
Am = AMist die gesamte Fläche der Membran und cm =<br />
cMist die flächenspezifische<br />
Membrankapazität.<br />
gK*(u^4)<br />
n^4<br />
(u-VK0)<br />
(Vm-Vk0)<br />
gNa*m^3<br />
*<br />
Product<br />
(u-VNa0)<br />
(Vm-VNa0)<br />
gkm<br />
(Vm-Vk0)<br />
gNam<br />
(Vm-VNa0)<br />
*<br />
Product<br />
*<br />
Product1<br />
1<br />
gkm*(Vm-Vk0)<br />
1<br />
gNam*(Vm-VNa0)
1<br />
Vm<br />
u-Vr<br />
V<br />
V<br />
(0.01*(10-u))/(exp((10-u)/10)-1)<br />
alpha_n<br />
0.125*exp(-u/80)<br />
beta_n<br />
1-u<br />
(1-n)<br />
alpha_*(1-n)<br />
<strong>Anhang</strong> 109<br />
<strong>Simulationsmodell</strong><br />
Abb. 82: Modul n(Vm). Berechnung des Faktors n in Abhängigkeit der Membranspannung<br />
VM() t und den empirischen Werten von Hodgkin und Huxley.<br />
1<br />
Vm<br />
(0.1*(25-u))/(exp((25-u)/10)-1)<br />
Abb. 83: Modul m(Vm). Berechnung des Faktors m in Abhängigkeit der Membranspannung<br />
VM() t und den empirischen Werten von Hodgkin und Huxley.<br />
1<br />
Vm<br />
u-Vr<br />
V<br />
u-Vr<br />
V<br />
Abb. 84: Modul h(Vm). Berechnung des Faktors h in Abhängigkeit der Membranspannung<br />
VM() t und den empirischen Werten von Hodgkin und Huxley.<br />
*<br />
*<br />
Product<br />
Product1<br />
beta_n*n<br />
alpha_m*(1-m)<br />
beta_m*m<br />
n<br />
Sum1<br />
n<br />
m<br />
m<br />
1<br />
s<br />
Integrator1<br />
Product<br />
alpha_m<br />
V<br />
1<br />
s<br />
1<br />
Sum1<br />
Integrator1<br />
4*exp(-u/18)<br />
beta_m<br />
0.07*exp(-u/20)<br />
1-u<br />
(1-m)<br />
*<br />
*<br />
Product1<br />
alpha_h*(1-h)<br />
Product<br />
alpha_h<br />
V<br />
1<br />
1<br />
s<br />
h<br />
Sum1<br />
Integrator1<br />
1/(exp((30-u)/10)+1)<br />
beta_h<br />
1-u<br />
(1-h)<br />
*<br />
*<br />
Product1<br />
beta_h*h<br />
h<br />
h<br />
1<br />
n
<strong>Anhang</strong> 110<br />
<strong>Simulationsmodell</strong><br />
Im unteren Teil der Abb. 79 wird der extrazelluläre Spannungsverlauf VJ() t im<br />
Spalt zwischen der Zellmembran und einer Oberfläche nach Gl.(54) berechnet. Der<br />
zeitliche Verlauf der Membranspannung VM() t wird aus der oberen Simulation übernommen.<br />
Die Module iK1 und iNa1 sind mit den Modulen iK und iNa identisch.<br />
An den Punkten mit der Bezeichnung Scope kann in der Software der zeitliche<br />
Verlauf des Signals ausgelesen werden.<br />
Tab. 9: Die Anfangswerte für die Simulation entsprechen dem Modell von<br />
Hudgkin und Huxley. Beim Einsetzen der Werte in das <strong>Simulationsmodell</strong><br />
können die Einheiten weggelassen werden. Die Zeitwerte haben dann die<br />
Einheit [ms].<br />
Am 3.1416 e-4 cm 2<br />
VK0 -62 mV<br />
VNa0 -39,4 mV<br />
Vr -50 mV<br />
gK 36 mS/cm 2<br />
gNa 120 mS/cm 2<br />
gL 0,3 mS/cm 2<br />
cm 1 μF/cm 2<br />
n0 0,3177<br />
m0 0,0529<br />
h0 0,5961
ii. Operationsverstärker<br />
ii.i. NF-Kenngrößen<br />
Differenzverstärkung<br />
Gleichtaktverstärkung<br />
Gleichtaktunterdrückung<br />
CMRR<br />
Eingangsruheströme<br />
<strong>Anhang</strong> 111<br />
Operationsverstärker<br />
Die Eigenschaften von Operationsverstärkern werden in den Datenblättern durch<br />
Kenngrößen beschrieben. Im folgenden Teil ii.i werden allgemein die Kenngrößen<br />
des Großsignalmodells und im Teil ii.ii die Kenngrößen für das Kleinsignalmodells<br />
eines Operationsverstärkers dargestellt. Für die durchgeführten Messungen wurde<br />
der Operationsverstärker vom Typ AD745 verwendet. Die wichtigsten Kenngrößen<br />
dieses Verstärkers werden in Abschnitt ii.iii vorgestellt.<br />
Am Ausgang des Operationsverstärkers stellt sich im unbeschalteten Zustand<br />
eine Spannung ein, die durch die Eingangsspannungsdifferenz zwischen nichtinvertierendem<br />
U + und invertierendem U - Eingang bestimmt wird.<br />
(172)<br />
Die Größe A d heißt Leerlauf-Differenzverstärkung (Open Loop Gain) und liegt<br />
normalerweise zwischen 10 3 und 10 6 . Damit läßt sich der Operationsverstärker als<br />
spannungsgesteuerte Spannungsquelle beschreiben. Der Eingangswiderstand wird<br />
als unendlich, der Ausgangswiderstand als Null angenommen.<br />
Bei einer Gleichtaktaussteuerung des Operationsverstärkers um ΔU gl , d.h. an beiden<br />
Eingängen liegt dieselbe Spannung an, müßte sich am Ausgang eine Spannung<br />
von Null Volt einstellen. Der reale Operationsverstärker zeigt aber eine<br />
Gleichtakteingangsspannungsabhängigkeit, die über die Gleichtaktverstärkung A gl<br />
ausgedrückt wird.<br />
(173)<br />
Das Verhältnis von Differenzverstärkung zur Gleichtaktverstärkung wird als<br />
Gleichtaktunterdrückung CMRR 1 bezeichnet.<br />
Die Ströme, die bei statischem Betrieb in die Eingänge eines realen Operationsverstärkers<br />
fließen, nennt man Eingangsruheströme. Bei bipolaren Operationsverstärkern<br />
entsprechen diese Ströme den Basiströmen der Eingangstransistoren.<br />
Operationsverstärker mit Feld-Effekt-Eingangstransistoren weisen aus diesem<br />
Grund typischerweise um drei Größenordnungen geringere Eingangsruheströme<br />
auf. Gleiche Ströme in beide Eingänge lassen sich durch schaltungstechnische<br />
Maßnahmen kompensieren.<br />
Offsetstrom Die Abweichung zwischen dem Eingangsstrom in den nichtinvertierenden und den<br />
invertierenden Eingang wird als Offsetstrom bezeichnet. Der Offsetstrom kann nicht<br />
kompensiert werden.<br />
Offsetspannung<br />
PSRR<br />
Ua = Ad U + – U- A gl<br />
( )<br />
ΔU a<br />
= ----------- =<br />
ΔU gl<br />
Ad ----------------<br />
CMRR<br />
Liegt an beiden Eingängen eine Spannung von Null Volt an, so ist bei einem realen<br />
Operationsverstärker die Ausgangsspannung von Null verschieden. Die Offsetspannung<br />
ist die Spannung, die an den Eingängen des Operationverstärkers<br />
angelegt werden muß, damit die Ausgangsspannung Null wird. Eine Änderung der<br />
Versorgungsspannung führt in der Regel auch zu einer Offsetspannungsänderung.<br />
Das Verhältnis der Offsetspannungsverschiebung zur Änderung der Versorgungsspannung<br />
wird als Betriebsspannungsdurchgriff PSRR 2 bezeichnet.<br />
1 Common Mode Rejection Ratio. Der Wert wird meist in dB angegeben.<br />
2 Power Supply Rejection Ratio. Der Wert wird meist in dB angegeben, wobei das Minuszeichen weggelassen<br />
wird.
ii.i.i. Großsignalbandbreite<br />
<strong>Anhang</strong> 112<br />
Operationsverstärker<br />
slew rate Kapazitive Effekte in der internen Schaltung des Operationsverstärkers begrenzen<br />
die maximale Anstiegsgeschwindigkeit oder Flankensteilheit (slew rate) S des<br />
Ausgangssignals. Sie wird in V/µs angegeben. Um eine unverzerrte Übertragung zu<br />
sichern, muß die Anstiegsgeschwindigkeit des Ausgangssignals kleiner sein als die<br />
maximale Flankensteilheit S. Für eine sinusförmige Ausgangsspannung der Amplitude<br />
û a<br />
folgt für die Änderungsgeschwindigkeit der Ausgangsspannung<br />
(174)<br />
dua -------- = ωûa cos(<br />
ωt)<br />
. (175)<br />
dt<br />
Ein Sinussignal mit der Frequenz f kann nur verzerrungsfrei übertragen werden,<br />
wenn gilt:<br />
S<br />
f < ----------- . (176)<br />
2ûaπ Diese Grenzfrequenz wird als Großsignalbandbreite bezeichnet. Sie ist in der<br />
Regel kleiner als die Grenzfrequenz, die sich aus dem Verstärkungs-Bandbreiten-<br />
Produkt ergibt.<br />
ii.ii.Kleinsignalmodell und Frequenzverhalten<br />
Eingangsimpendanzen<br />
ua() t = ûa sin(<br />
ωt)<br />
Ein vereinfachtes Kleinsignalersatzschaltbild eines Operationsverstärkers ist in<br />
Abb. 85 dargestellt. Die Gleichtakt- und Differenzeingangsimpedanzen werden in<br />
den Datenblättern jeweils durch die Werte eines Widerstandes r gl bzw. r d und einer<br />
Kapazität c gl bzw. c d angegeben, die zueinander parallelgeschaltet sind und für<br />
reine Gleichtakt- bzw. Differenzaussteuerung gelten. Für die Elemente des Ersatzschaltbildes<br />
gelten folgende Beziehungen:<br />
rgl ′ = 2rgl cgl ′<br />
=<br />
cgl ------<br />
2<br />
(177)<br />
rd cgl rd ′ = ------------------<br />
cd ′ = cd – ------ . (178)<br />
rd 4<br />
1 – ---------<br />
4r gl<br />
Der Frequenzgang der Differenz- und Gleichtaktverstärkung wird im Ersatzschaltbild<br />
nicht durch ein Netzwerk nachgebildet, sondern in frequenzabhängigen Übertragungsfaktoren<br />
ad bzw. agl berücksichtigt. Die Summe der Spannungen am<br />
Ausgang ergibt die Leerlaufspannung. Die Lastabhängigkeit der Ausgangsspannung<br />
wird durch ra berücksichtigt.
Tiefpaßcharakteristik<br />
u +<br />
u -<br />
u d<br />
Abb. 85: Kleinsignalersatzschaltbild eines Operationsverstärkers<br />
<strong>Anhang</strong> 113<br />
Operationsverstärker<br />
Die Differenzverstärkung der Operationsverstärker weist eine Tiefpaßcharakteristik<br />
höherer Ordnung auf. Sie kann durch die Hintereinanderschaltung von spannungsgesteuerten<br />
Spannungsquellen mit Übertragungsfaktor „1“ und einer der<br />
Grenzfrequenz fc entsprechenden RC-Kombination nachgebildet werden. Für den<br />
Übertragungsfaktor der Differenzverstärkung ad mit einem Tiefpaßverhalten 1. Ordnung<br />
gilt:<br />
a d<br />
A d<br />
mit (179)<br />
1 j f<br />
1<br />
= -----------------<br />
fc0 = --------------<br />
+ -----<br />
2πRC<br />
f c0<br />
Für f → 0 gilt ad → Ad und agl →<br />
Agl .<br />
r' d<br />
c' d<br />
Abb. 86: Frequenzgang von Betrag und Phasenverschiebung der Leerlaufdifferenzverstärkung<br />
sowie CMRR [5]<br />
Bandbreite Die Frequenz f co stellt die 3dB-Knick-Frequenz des Amplitudengangs der Differenzverstärkung<br />
dar und entspricht beim nichtrückgekoppelten Operationsverstärker<br />
der Bandbreite. Für Frequenzen f>f c0 fällt |a d | mit 20dB / Dekade ab.<br />
Transitfrequenz Die Transitfrequenz f T kennzeichnet die Frequenz, bei der die Differenzverstärkung<br />
auf 0dB abgefallen ist.<br />
2r gl<br />
2r gl<br />
c/2<br />
gl<br />
c/2<br />
gl<br />
au au<br />
d d gl gl<br />
r a
ii.iii.Operationsverstärker Typ AD745<br />
<strong>Anhang</strong> 114<br />
Operationsverstärker<br />
Der Operationsverstärker AD745 besitzt einen FET-Eingang, so daß nur sehr<br />
kleine Eingangsruheströme fließen. Gleichzeitig zeichnet er sich durch sehr geringes<br />
Rauschen und ein hohes Gain-Bandwidth-Produkt aus. Die Tab. 10 enthält die<br />
wichtigsten Kenngrößen in einer Übersicht, weitere Daten sind im Datenblatt enthalten<br />
(<strong>Anhang</strong> CD-ROM).<br />
Tab. 10:Kenngrößen AD745<br />
Open Loop Gain<br />
(Differenzverstärkung)<br />
CMRR<br />
(Gleichtaktunterdrückung)<br />
Input Bias Current<br />
(Eingangsruheströme)<br />
Input Offset Current<br />
(Offsetstrom)<br />
1000-4000<br />
80-95<br />
150-400<br />
40-150<br />
PSRR 90-96<br />
Slew Rate (Gain = -4) 12,5<br />
Gain Bandwidth Product 20<br />
Input Current Noise (1kHz) 6,9<br />
Input Voltage Noise (10kHz) 2,9<br />
V/mV<br />
dB<br />
pA<br />
pA<br />
dB<br />
V/μs<br />
MHz<br />
fA/ Hz<br />
nV/ Hz
iii.Schaltungsaufbau<br />
iii.i.Multiplexer<br />
<strong>Anhang</strong> 115<br />
Schaltungsaufbau<br />
Die Abb. 87 zeigt die Platine des Multiplexers und der IU-Umsetzer von oben, die<br />
im Rahmen dieser Arbeit hergestellt wurde. Die linke Reihe der blauen DIP-Schalter<br />
dient zur Auswahl der Drain-Zuleitungen für die linken Transistoren, die rechte<br />
Reihe zur Auswahl der Drain-Zuleitungen für die rechten Transistoren. Mit den<br />
unteren beiden DIP-Schaltern der mittleren Reihe werden die Drain-Zuleitungen mit<br />
einem der vier Ausgänge (Drain extern 1, Drain extern 2, IU-Wandler 1, IU-Wandler<br />
2) verbunden.<br />
Abb. 87: Aufsicht auf die Platine des Multiplexers und der IU-Umsetzer.
iii.ii.IU-Umsetzer<br />
<strong>Anhang</strong> 116<br />
Schaltungsaufbau<br />
Die oberen beiden DIP-Schalter der mittleren Reihe dienen zur Auswahl der<br />
Source-Zuleitungen der Transistoren. Die DIP-Schalter haben drei Stellungen<br />
(oben: ein, mitte: hochohmig, unten: Masse). Mit den roten DIP-Schaltern können<br />
die zweiten Drain-Zuleitungen der Transistoren „d“ zugeschaltet werden.<br />
Auf dem unteren Drittel der Platine befinden sich zwei IU-Umsetzer, die fast symmetrisch<br />
aufgebaut sind (rechts und links unten). Sie unterscheiden sich nur durch<br />
unterschiedliche Operationsverstärkerbausteine in der ersten Stufe.<br />
Abb. 88: Platine (unten). Im endgültigen Platinenlayout sind alle freien Flächen<br />
als Masseflächen ausgeführt. (Beschriftung fehlt noch)
Abb. 89: Platine (oben).<br />
<strong>Anhang</strong> 117<br />
Schaltungsaufbau
iv.Kennlinien der EOS-Transistoren<br />
<strong>Anhang</strong> 118<br />
Kennlinien der EOS-Transistoren<br />
Im folgenden sind die Kennlinien der 16 unterschiedlichen EOS-Transistoren auf<br />
dem Cultus-Chip dargestellt. Die Messungen wurden jeweils am Transistor „a“ in<br />
der entsprechenden Reihe (1 bis 16) durchgeführt.<br />
Aus den Ausgangskennlinien mit UDS = UGS (Teil iv.i) werden die Übertragungsleitwertfaktoren<br />
β und die Einsatzspannungen UTH bestimmt. Die genaue Vorgehensweise<br />
ist in Kap. 5.1 beschrieben. Aus den Steuerkennlinien (Transferstromkennlinie)<br />
mit UDS = – 25V , (Teil iv.ii) wird die Steilheit der Transistoren gm im<br />
gewählten Arbeitspunkt abgelesen. Die Tab. 11 enthält zur Übersicht alle ermittelten<br />
Parameter.<br />
Tab. 11:Parameter der EOS-Transistoren. Die Steilheit gm bezieht sich auf den<br />
Arbeitspunkt UDS = UGS = – 25V , .<br />
Transistor W/L β [10 -6 A/V 2 ] K p [10 -6 A/V 2 ] U TH [V] g m [µS]<br />
1 4 239 59,75 -0,29 340,7<br />
2 6 394 65,67 -0,32 486,5<br />
3 3 183 61 -0,32 255,8<br />
4 2 110 55 -0,33 161,3<br />
5 2 72 36 -0,32 121,1<br />
6 1,33 60 45,11 -0,35 101,3<br />
7 2,67 143 53,56 -0,31 222,7<br />
8 1,11 48 43,24 -0,49 78,2<br />
9 1 50 50 -0,32 89,9<br />
10 1 47 47 -0,32 79,3<br />
11 1 51 51 -0,33 85,2<br />
12 1,11 47 42,34 -0,40 75,8<br />
13 1,11 27 27 -0,47 49,2<br />
14 1 56 56 -0,35 92,5<br />
15 0,2 7 35 -0,33 14,3<br />
16 5 391,7<br />
iv.i.Ausgangskennlinien mit U GS =U DS<br />
In den Abb. 90 bis Abb. 119 ist für jeden Transistortyp des Cultus-Chips die Ausgangskennlinie<br />
IDS = fU ( DS)<br />
und IDS = fU ( DS)<br />
mit UDS = UGS dargestellt.<br />
Die Kennlinie IDS = fU ( DS)<br />
ist im Idealfall eine Gerade mit der Steigung β/2 ,<br />
sofern sich der Transistor immer im Sättigungsbereich befindet. Dies ist der Fall,<br />
wenn UDS = UGS gilt. Aus dem Schnittpunkt der Geraden mit dem Nullpunkt kann<br />
die Einsatzspannung UTH und aus der Steigung der Geraden kann der Übertragungsleitwertfaktor<br />
β<br />
bestimmt werden (Kap. 5.1).
-I ds [A]<br />
800µ<br />
700µ<br />
600µ<br />
500µ<br />
400µ<br />
300µ<br />
200µ<br />
100µ<br />
0<br />
Abb. 90: Transitor 1: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
30m<br />
25m<br />
20m<br />
15m<br />
10m<br />
5m<br />
0<br />
– 6<br />
Abb. 91: Transistor 1: = – 029V , , β 239×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 119<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
1m<br />
800µ<br />
600µ<br />
400µ<br />
200µ<br />
0<br />
Abb. 92: Transistor 2: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
35m<br />
30m<br />
25m<br />
20m<br />
15m<br />
10m<br />
5m<br />
0<br />
– 6<br />
Abb. 93: Transistor 2: = – 032V , , β 394×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 120<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
500µ<br />
400µ<br />
300µ<br />
200µ<br />
100µ<br />
0<br />
Abb. 94: Transistor 3: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
25m<br />
20m<br />
15m<br />
10m<br />
5m<br />
0<br />
– 6<br />
Abb. 95: Transistor 3: = – 032V , , β 183×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 121<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
350µ<br />
300µ<br />
250µ<br />
200µ<br />
150µ<br />
100µ<br />
50µ<br />
0<br />
Abb. 96: Transistor 4: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
20m<br />
15m<br />
10m<br />
5m<br />
0<br />
– 6<br />
Abb. 97: Transistor 4: = – 033V , , β 110×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 122<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
250µ<br />
200µ<br />
150µ<br />
100µ<br />
50µ<br />
0<br />
Abb. 98: Transistor 5: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
16m<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 99: Transistor 5: = – 032V , , β 72×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 123<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
250µ<br />
200µ<br />
150µ<br />
100µ<br />
50µ<br />
0<br />
Abb. 100:Transistor 6: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
16m<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 101:Transistor 6: = – 035V , , β 60×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 124<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
450µ<br />
400µ<br />
350µ<br />
300µ<br />
250µ<br />
200µ<br />
150µ<br />
100µ<br />
50µ<br />
0<br />
Abb. 102:Transistor 7: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
20m<br />
15m<br />
10m<br />
5m<br />
0<br />
– 6<br />
Abb. 103:Transistor 7: = – 031V , , β 143×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 125<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS ]
I D [A]<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
0<br />
Abb. 104:Transistor 8: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 105:Transistor 8: = – 049V , , β 48×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 126<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
180µ<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
0<br />
Abb. 106:Transistor 9: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 107:Transistor 9: = – 032V , , β 50×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 127<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =UGS)
-I DS [A]<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
0<br />
Abb. 108:Transistor 10: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 109:Transistor 10: = – 032V , , β 47×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 128<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
0<br />
Abb. 110:Transistor 11: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [ sqrt(A)]<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 111:Transistor 11: = – 033V , , β 51×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 129<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
0<br />
Abb. 112:Transistor 12: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 113:Transistor 12: = – 040V , , β 47×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 130<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
100µ<br />
90µ<br />
80µ<br />
70µ<br />
60µ<br />
50µ<br />
40µ<br />
30µ<br />
20µ<br />
10µ<br />
0<br />
Abb. 114:Transistor 13: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 115:Transistor 13: = – 047V , , β 27×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 131<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
180µ<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
0<br />
Abb. 116:Transistor 14: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
14m<br />
12m<br />
10m<br />
8m<br />
6m<br />
4m<br />
2m<br />
0<br />
– 6<br />
Abb. 117:Transistor 14: = – 035V , , β 56×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 132<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
-I DS [A]<br />
30µ<br />
25µ<br />
20µ<br />
15µ<br />
10µ<br />
5µ<br />
0<br />
Abb. 118:Transistor 15: Ausgangskennlinie mit = .<br />
sqrt(-I DS ) [sqrt(A)]<br />
5m<br />
4m<br />
3m<br />
2m<br />
1m<br />
0<br />
– 6<br />
Abb. 119:Transistor 15: = – 033V , , β 7×10<br />
A/V .<br />
2<br />
=<br />
<strong>Anhang</strong> 133<br />
Kennlinien der EOS-Transistoren<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
-U DS [V] (U DS =U GS )<br />
U DS<br />
U GS<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U TH<br />
-U DS [V] (U DS =U GS )
iv.ii.Stromsteuerkennlinien<br />
-I DS [A]<br />
-I DS [A]<br />
700µ<br />
600µ<br />
500µ<br />
400µ<br />
300µ<br />
200µ<br />
100µ<br />
Abb. 120:Transistor 1: = – 25V , , gm = 340, 7μS<br />
800µ<br />
600µ<br />
400µ<br />
200µ<br />
Abb. 121:Transistor 2: = – 25V , , gm =<br />
486, 5μS<br />
<strong>Anhang</strong> 134<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
1m<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
-I DS [A]<br />
-I DS [A]<br />
500µ<br />
400µ<br />
300µ<br />
200µ<br />
100µ<br />
Abb. 122:Transistor 3: = – 25V , , gm = 255, 8μS<br />
350µ<br />
300µ<br />
250µ<br />
200µ<br />
150µ<br />
100µ<br />
50µ<br />
Abb. 123:Transistor 4: = – 25V , , gm =<br />
161, 3μS<br />
<strong>Anhang</strong> 135<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
-I DS [A]<br />
-I DS [A]<br />
250µ<br />
200µ<br />
150µ<br />
100µ<br />
50µ<br />
Abb. 124:Transistor 5: = – 25V , , gm = 121, 1μS<br />
200µ<br />
180µ<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
Abb. 125:Transistor 6: = – 25V , , gm =<br />
101, 3μS<br />
<strong>Anhang</strong> 136<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
-I DS [A]<br />
-I DS [A]<br />
450µ<br />
400µ<br />
350µ<br />
300µ<br />
250µ<br />
200µ<br />
150µ<br />
100µ<br />
50µ<br />
Abb. 126:Transistor 7: = – 25V , , gm = 222, 7μS<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
Abb. 127:Transistor 8: = – 25V , , gm =<br />
78, 2μS<br />
<strong>Anhang</strong> 137<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
-I DS [A]<br />
-I DS [A]<br />
180µ<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
Abb. 128:Transistor 9: = – 25V , , gm = 89, 9μS<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
Abb. 129:Transistor 10: = – 25V , , gm =<br />
79, 3μS<br />
<strong>Anhang</strong> 138<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
-I DS [A]<br />
-I DS [A]<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
Abb. 130:Transistor 11: = – 25V , , gm = 85, 2μS<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
Abb. 131:Transistor 12: = – 25V , , gm =<br />
75, 8μS<br />
<strong>Anhang</strong> 139<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
-I DS [A]<br />
-I DS [A]<br />
100µ<br />
90µ<br />
80µ<br />
70µ<br />
60µ<br />
50µ<br />
40µ<br />
30µ<br />
20µ<br />
10µ<br />
Abb. 132:Transistor 13: = – 25V , , gm = 49, 2μS<br />
180µ<br />
160µ<br />
140µ<br />
120µ<br />
100µ<br />
80µ<br />
60µ<br />
40µ<br />
20µ<br />
Abb. 133:Transistor 14: = – 25V , , gm =<br />
92, 5μS<br />
<strong>Anhang</strong> 140<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
-I DS [A]<br />
-I DS [A]<br />
30µ<br />
25µ<br />
20µ<br />
15µ<br />
10µ<br />
5µ<br />
Abb. 134:Transistor 15: = – 25V , , gm = 14, 3μS<br />
800µ<br />
600µ<br />
400µ<br />
200µ<br />
Abb. 135:Transistor 16: = – 25V , , gm =<br />
391, 7μS<br />
<strong>Anhang</strong> 141<br />
Kennlinien der EOS-Transistoren<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)<br />
0<br />
0,0 0,5 1,0 1,5 2,0 2,5 3,0<br />
U DS<br />
-U GS [V] (U DS =-2,5V)
v. Präparation von Herzmuskelzellen<br />
Ansetzen der<br />
Lösung<br />
<strong>Anhang</strong> 142<br />
Präparation von Herzmuskelzellen<br />
Zur Präparation von Herzmuskelzellen aus erwachsenen Ratten (adult) werden<br />
folgende Lösungen 1 benötigt: Ca ++ -freie-Lösung (Tab. 12), Ca ++ -10mM-Lösung<br />
(Tab. 13), Extrazelluläre Lösung (Tab. 14). Zum Aufbewahren von Herzmuskelzellen<br />
wird die Glutamat KB Lösung (Tab. 16) verwendet, zum Patchen der Zellen muß<br />
zusätzlich die intrazelluläre Lösung (Tab. 15) angesetzt werden.<br />
Tab. 12:Ca ++ -freie-Lösung (pH 7,1 mit NaOH). Die Glucose wird erst kurz vor der<br />
Präparation hinzugegeben (MGW: Masse pro Mol).<br />
MGW Konzentration<br />
[mM]<br />
Damit die Ca ++ -freie-Lösung (Tab. 12) längere Zeit im Kühlschrank aufbewahrt<br />
werden kann, wird die Glucose zunächst weggelassen. Erst kurz vor Verwendung<br />
wird der entsprechende Anteil an Glucose gelöst. (In 150ml Ca ++ -freie-Lösung müssen<br />
0,300g Glucose gelöst werden.)<br />
Zur Herstellung der Ca ++ -freien-Lösung werden die Substanzen abgewogen und<br />
in destilliertem Wasser gelöst. Es wird eine um ca. 10% geringere Menge destilliertes<br />
Wasser als die angestrebte Lösungsmenge verwendet. Zur Einstellung des pH-<br />
Wertes wird unter ständigem Rühren tropfenweise gelöstes Natriumhydroxid (1M<br />
NaOH) hinzugegeben, bis der gewünschte pH-Wert erreicht ist. Die Lösung wird<br />
anschließend mit destilliertem Wasser bis zur angestrebten Menge aufgefüllt. Diese<br />
Vorgehensweise ist möglich, da die Lösung durch Hepes gepuffert wird. Die weiteren<br />
Lösungen werden analog angesetzt.<br />
Tab. 13:Ca ++ 10mM Lösung (CaCl 2 Lösung)<br />
Lösung<br />
1000 ml<br />
Lösung<br />
2000 ml<br />
NaCl 58,4 g 140,0 8,182 g 16,364 g<br />
KCl 74,6 g 5,8 0,432 g 0,864 g<br />
KH 2PO 4 136,0 g 0,5 0,068 g 0,136 g<br />
Na 2HPO 4<br />
(Na 2 HPO 4 2H 2 O)<br />
142,5 g<br />
177,9 g<br />
0,4<br />
0,4<br />
0,057 g<br />
0,071 g<br />
0,114 g<br />
0,142 g<br />
MgSo 4 120,0 g 0,9 0,108 g 0,216 g<br />
Glucose 180,2 g 11,1 2,000 g 4,000 g<br />
Hepes 238,3 g 10,0 2,383 g 4,766 g<br />
MGW Konzentration<br />
[mM]<br />
Lösung<br />
100 ml<br />
Lösung<br />
50 ml<br />
CaCl 2 2H 2 O 147,0 g 10 0,147 g 0,073 g<br />
1 Eine 1-molare Lösung (1 M Lösung) enthält immer 1 mol der betreffenden Substanz in 1 Liter Lösung.
Tab. 14:Extrazelluläre Lösung (pH 7,3 mit NaOH)<br />
Tab. 15:Intrazelluläre Lösung (pH 7,3 mit KOH)<br />
<strong>Anhang</strong> 143<br />
Präparation von Herzmuskelzellen<br />
Tab. 16:Glutamat KB (pH 7,3 mit KOH). Das Glutamat KB kann in kleinen<br />
Portionen eingefroren und längere Zeit aufbewahrt werden.<br />
v.i. Vorbereitung der Präparation<br />
Zur Präparation der Herzmuskelzellen werden neben den oben beschriebenen<br />
Lösungen folgende Materialien benötigt:<br />
5 Bechergläser 30ml<br />
1 Becherglas 300ml<br />
1 Becherglas 500ml<br />
1 Petrischale<br />
2 Spritzen 10ml<br />
2 Spritzen 5ml<br />
1 Glaspipette<br />
Einhängeglas<br />
MGW Konzentration<br />
[mM]<br />
Lösung<br />
1000 ml<br />
Lösung<br />
500 ml<br />
NaCl 58,4 g 150,0 8,766 g 4,383 g<br />
KCl 74,6 g 5,4 0,403 g 0,201 g<br />
Hepes 238,3 g 5,0 1,191 g 0,596 g<br />
CaCl 2 147,0 g 2,5 0,367 g 0,184 g<br />
MgCl 2 203,3 g 0,5 0,102 g 0,051 g<br />
MGW Konzentration<br />
[mM]<br />
Lösung<br />
1000 ml<br />
Lösung<br />
100 ml<br />
KCl 74,6 g 150 11,184 g 1,118 g<br />
Hepes 238,3 g 5 1,191 g 0,119 g<br />
EGTA 380,4 g 10 3,804 g 0,3804<br />
MGW Konzentration<br />
[mM]<br />
Lösung<br />
500 ml<br />
Lösung<br />
1000 ml<br />
Glutaminsäure 147,2 g 50,0 3,680 g 7,216 g<br />
HEPES 238,5 g 20,0 2,385 g 4,770 g<br />
Taurin 125,0 g 20,0 1,250 g 2,500 g<br />
Glucose 180,0 g 10,0 0,900 g 1,800 g<br />
MgSO 4 120,0 g 3,0 0,180 g 0,360 g<br />
EGTA 380,0 g 0,5 0,095 g 0,190 g<br />
KCl 74,6 g 30,0 1,118 g 2,236 g<br />
KH 2 PO 4 136,1 g 30,0 2,042 g 4,084 g<br />
KOH Plätzchen 10 Stück 20 Stück
einstellbare Pipette 100μl<br />
Kanüle mit Drei-Wege-Hahn<br />
Faden<br />
Hirschmannklemme<br />
Eis<br />
Präparationsbesteck.<br />
<strong>Anhang</strong> 144<br />
Präparation von Herzmuskelzellen<br />
Thermostat auf 37°C vorheizen und eingefrohrene Glutamat KB Lösung im Wasserbad<br />
auftauen. Zur Ca ++ -freien-Lösung Glucose hinzugeben. In 150ml Ca ++ freier<br />
Lösung müssen 0,300g Glucose gelöst werden. Die Ca ++ -freie-Lösung sollte zum<br />
Kühlen auf Eis aufbewahrt werden. 5mg Enzym (Kollagenase A, Bochinger Mannheim<br />
GmbH) abwiegen, in 15ml Ca ++ freier Lösung (mit Glucose) auflösen und 15μl<br />
Ca ++ 10mM hinzugeben.<br />
Die Apparatur sollte vor jeder Präparation sehr gut mit destilliertem Wasser<br />
gespült werden. In Intervallen von 3-4 Präparationen ist es sinnvoll, die Apparatur<br />
mit Wasserstoffperoxid (6%) und Ethanol (100%) zu reinigen. Zum Füllen der Apparatur<br />
eine 10ml Spritze mit Ca ++ -freier-Lösung füllen und an den Drei-Wege-Hahn 2<br />
(Abb. 136) ansetzen. Zunächst die Leitung zum Enzymgefäß, dann die Leitung zum<br />
Gefäß für die Ca ++ -freie-Lösung füllen. Es sollte darauf geachtet werden, daß die<br />
Leitungen blasenfrei sind. Gefäß für Ca ++ -freie-Lösung füllen, Enzymgefäß füllen<br />
und restliche Enzym-Lösung in das Einhängeglas geben. 5ml Spritze mit Drei-<br />
Wege-Hahn und Kanüle mit Ca ++ -freier-Lösung blasenfrei füllen. Fäden, Hirschmannklemme<br />
und Petrischale bereitlegen. In die Petrischale wird kurz vor Beginn<br />
der Präparation etwas Ca ++ -freie-Lösung gegossen.<br />
Präparationsplatz mit Papiertüchern auslegen und 30ml Becherglas mit Ca ++ -<br />
freier-Lösung für den Transport des Herzens füllen.<br />
Einhängeglas<br />
Enzym Ca ++ freie Lösung<br />
4 5<br />
Doppelwandgefäß<br />
Thermostat<br />
Kanüle<br />
Thermostat<br />
Abb. 136:Apparatur zur Präparation von Herzmuskelzellen<br />
3<br />
2<br />
1
v.ii.Durchführung<br />
<strong>Anhang</strong> 145<br />
Präparation von Herzmuskelzellen<br />
Nach der Betäubung der Ratte muß möglichst schnell das Herz mit einem langen<br />
Aortaansatz herauspräpariert werden. Es ist besonders darauf zu achten, daß das<br />
Herz nicht beschädigt wird. Das Herz wird in Ca ++ -freier-Lösung in die Petrischale<br />
gelegt. Vorsichtig werden die Lungenreste und anderes Gewebe entfernt und die<br />
Aorta freigelegt. Die Verzweigungen an der Aorta werden ggf. abgeschnitten, um<br />
die Kanüle einzuführen. Dabei darf die Kanüle nicht zu weit in das Herz eingeführt<br />
werden. Die Aorta wird zunächst mit der Hirschmannklemme an der Kanüle fixiert<br />
und anschließend mit einem dünnen Faden festgebunden. Damit die Kanüle nicht<br />
aus der Aorta herausrutschen kann, ist auf der Spitze der Kanüle ein Stück Gummischlauch<br />
aufgeschoben und mit Sekundenkleber fixiert. Durch leichten Druck auf<br />
die Spritze werden die Herzkranzgefäße und die Herzkammern freigespült. Erst<br />
nach erfolgreichem Freispülen wird das Herz mit dem Drei-Wege-Hahn an der<br />
Apparatur befestigt. Bevor der Drei-Wege-Hahn 1 geöffnet wird, muß mit einer<br />
Spritze die Luft zwischen Drei-Wege-Hahn 1 und 2 herausgedrückt werden. Das<br />
Herz wird 5 Minuten lang mit Ca ++ -freier-Lösung ausgewaschen. Dabei sollte das<br />
Herz massiert werden, um die Blutreste möglichst vollständig zu entfernen. Die<br />
Spülung erfolgt über einem Becherglas.<br />
Nach 5 Minuten wird auf Enzymspülung umgeschaltet. Das Einhängeglas wird so<br />
unter das Herz geschwenkt, daß das Herz in der Enzym-Lösung hängt. Mit einer<br />
Glaspipette wird laufend Enzym-Lösung vom Einhängegefäß in das obere Gefäß<br />
zurückgefüllt. Nach Ablauf der ersten 4 Minuten werden 15μl Ca ++ -10mM dazugegeben,<br />
nach allen weiteren 4 Minuten werden jeweils 30μl Ca ++ -10mM hinzugegeben.<br />
Insgesamt wird das Herz 24 Minuten mit Enzym-Lösung gespült. Vor dem<br />
Freispülen des Herzens vom Enzym wird die restliche Ca ++ -freie-Lösung aus der<br />
Apparatur von unten herausgesaugt. Mit einer Spritze werden anschließend von<br />
unten 5ml Glutamat KB Lösung eingefüllt. Zum vollständigen Freispülen muß ggf.<br />
Druck auf Hahn 5 angelegt werden, damit in ca. 5 min die gesamte Lösung durchgelaufen<br />
ist.<br />
Das Herz wird im oberen Drittel von der Kanüle abgeschnitten und über kreuz<br />
zweimal von der Schnittkante her eingeschnitten. Zwei Bechergläser werden mit<br />
etwas Glutamat KB Lösung gefüllt und das Herz wird im ersten Becherglas ca. 30<br />
Sekunden, im zweiten Becherglas ca. 60 Sekunden geschwenkt. Jeweils ca. 2ml<br />
der Zellemulsion werden auf eine 10ml Ampulle mit Glutamat KB Lösung verdünnt.<br />
Abschließend muß die Apparatur mit destilliertem Wasser gründlich gespült werden.
vi.Patch-Anleitung<br />
<strong>Anhang</strong> 146<br />
Patch-Anleitung<br />
(1) Chip in Sockel einsetzen. Erdung, Flow-Zu- und Abfluß justieren. Schale mit<br />
extrazellulärer Lösung füllen.<br />
(2) Chip mit Objektiv 1 scharfstellen.<br />
(3) Mit Pipette ein bis zwei Tropfen Zellemulsion auf den Chip tropfen und Zellen<br />
absetzen lassen.<br />
(4) Schmutz und Kraftbrühe mit mind. 10ml extrazellulärer Lösung wegspülen.<br />
Die Kraftbrühe verhindert einen guten Seal.<br />
(5) Zelle aussuchen: Am besten eine Zelle wählen, die direkt auf einem Transistor<br />
liegt. Sonst möglichst eine Zelle aussuchen, die sich beim Spülen noch<br />
lange verschoben hat. Langsames Zucken der Zelle ist unproblematisch. Die<br />
Zelloberfläche sollte keine sehr stark ausgeprägte „Maiskolben-Struktur“<br />
haben.<br />
(6) Zelle mit Objektiv 2 und anschließend mit Objektiv 3 scharfstellen.<br />
(7) Pipette mit intrazellulärer Lösung füllen. Erst von unten über Kapillarwirkung,<br />
dann mit Kanüle von oben, Luftblasen herausklopfen.<br />
(8) Flow-Zu- und Abfluß entfernen.<br />
(9) Pipette einsetzen, mit Objektiv 1 scharfstellen und senkrecht ausrichten.<br />
(10) Überdruck auf Pipette geben (ca. 15cm Wassersäule)<br />
(11) Patch-Clamp-Verstärker auf VC-Search stellen. Testimpuls (10ms, 10mV)<br />
einstellen.<br />
(12) Pipette in Elektrolyten eintauchen.<br />
(13) Pipettenwiderstand bestimmen. Offset abgleichen. Gute Pipetten für Herzmuskelzellen<br />
liegen zwischen 1,7 und 2,2MΩ; auf VC umstellen.<br />
(14) Pipettenschaft mit Objektiv 1 und anschließend mit Objektiv 2 Pipettenöffnung<br />
scharfstellen.<br />
(15) Pipette absenken und Schärfe nachstellen.<br />
(16) Zu Objektiv 3 wechseln und Pipette weiter absenken.<br />
(17) Pipette so weit auf die Membran der Zelle absenken bis sie sich deutlich eindellt<br />
und der Widerstand um 3-4MΩ angestiegen ist. Letztes Heranfahren mit<br />
langsamster Geschwindigkeit.<br />
(18) Druck ablassen und ca. 10cm Wassersäule Unterdruck geben.<br />
(19) Sealen abwarten. Verstärkung nachstellen (am Verstärker und in der Software).<br />
(20) Ab 100MΩ Haltespannung langsam verkleinern.<br />
(21) Vor dem Durchreißen der Membran muß die Haltespannung bei -80mV liegen.<br />
(22) Durchreißen der Membran: Ohne Wassersäule 5ml Spritze durchziehen.<br />
Wenn die Membran durchreißt, übersteuert der Verstärker. Verstärkung auf<br />
Gain 1 zurückstellen und auf CC-fast umschalten. Protokoll zu CC wechseln.<br />
(23) Wenn der Seal gut ist, wird die Zelle bei -70 bis -80mV geklemmt, sonst<br />
leicht negativen Haltestrom einstellen.<br />
(24) Stimulationsimpuls geben. Stromimpuls schrittweise erhöhen, bis Aktionspotential<br />
ausgelöst wird. Dann Impulslänge verkleinern.<br />
(25) Transistor auswählen: Liegt die Zelle schon auf dem richtigen Transistor,<br />
muß unbedingt der Transistor vor dem Patchen eingeschaltet werden, sonst<br />
wird die Zelle durch Wackeln zerstört. Muß die Zelle umgesetzt werden,<br />
Transistor einschalten, sobald die Zelle angehoben ist.<br />
(26) Umsetzen der Zellen: Pipette mit langsamster Geschwindigkeit anheben,<br />
dabei leicht diagonal nach oben ziehen; dann Chip unter der Pipette verschieben.<br />
Zelle über Transistor absenken.<br />
(27) Bei den Messungen muß das Licht ausgeschaltet werden (Fotoeffekt).
<strong>Anhang</strong> 147<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
vii.Ergebnisse aus Messungen mit EOS-<br />
Transistoren an Herzmuskelzellen<br />
Bei den durchgeführten Messungen wurden die Herzmuskelzellen über eine<br />
Patch-Pipette periodisch stimuliert und der intrazelluläre Potentialverlauf registriert.<br />
Zeitgleich wurde die Modulation des Source-Drain-Stroms des Transistors unterhalb<br />
der Zelle in eine Spannung umgesetzt und verstärkt. Der intrazelluläre Potentialverlauf,<br />
die Modulation des Transistorstroms und das Stimulationssignal wurden<br />
mit der Patch-Clamp-Software (ISO2) aufgezeichnet.<br />
Die Daten der Messung an Zelle 7 über Transistor 5b sind in der Datei<br />
„051099id.3“ und die Daten der Messung an Zelle 16 über Transistor 5c sind in der<br />
Datei „071099id.3“ enthalten.<br />
In den folgenden Diagrammen der Abb. 137 bis Abb. 150 sind Mittlungen der<br />
Signale zu unterschiedlichen Zeitpunkten der Messungen und über eine unterschiedliche<br />
Anzahl von Perioden dargestellt.
Strommodulation Transistor [A]<br />
intracelluläre Spannung [mV]<br />
Stimulus [A]<br />
1n<br />
0<br />
60<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
200p<br />
0<br />
-200p<br />
-400p<br />
<strong>Anhang</strong> 148<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 137:Messung an Zelle 7 mit Transistor 5b. Mittelwert über 91 Messungen<br />
(Bilder 550-640 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
intracelluläre Spannung [mV]<br />
Stimulus [A]<br />
1n<br />
0<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
<strong>Anhang</strong> 149<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
-1n<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 138:Messung an Zelle 7 mit Transistor 5b. Mittelwert über 50 Messungen<br />
(Bilder 725-774 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
intracelluläre Spannung [mV]<br />
Stimulus [A]<br />
1n<br />
0<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
<strong>Anhang</strong> 150<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 139:Messung an Zelle 7 mit Transistor 5b. Mittelwert über 27 Messungen<br />
(Bilder 786-812 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
intracelluläre Spannung [mV]<br />
Stimulus [A]<br />
1n<br />
0<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
2n<br />
1n<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 151<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 140:Messung an Zelle 7 mit Transistor 5b. Mittelwert über 7 Messungen<br />
(Bilder 813-819 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 152<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 141:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 39 Messungen<br />
(Bilder 061-099 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
1n<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 153<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 142:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 9 Messungen<br />
(Bilder 101-109 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 154<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 143:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 73 Messungen<br />
(Bilder 101-173 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 155<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 144:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 10 Messungen<br />
(Bilder 155-164 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 156<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 145:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 61 Messungen<br />
(Bilder 184-244 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 157<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
-2n<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 146:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 162 Messungen<br />
(Bilder 245-407 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
40<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
-80<br />
500p<br />
0<br />
-500p<br />
-1n<br />
<strong>Anhang</strong> 158<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
-2n<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 147:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 104 Messungen<br />
(Bilder 409-512 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
20<br />
0<br />
-20<br />
-40<br />
-60<br />
<strong>Anhang</strong> 159<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
-80<br />
10 20 30 40 50 60 70<br />
1n<br />
500p<br />
0<br />
-500p<br />
-1n<br />
time [ms]<br />
-2n<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 148:Messung an Zelle 16 mit Transistor 5c. Mittelwert über 10 Messungen<br />
(Bilder 503-512 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
20<br />
10<br />
0<br />
-10<br />
<strong>Anhang</strong> 160<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
-20<br />
10 20 30 40 50 60 70<br />
2n<br />
0<br />
-2n<br />
-4n<br />
-6n<br />
-8n<br />
-10n<br />
time [ms]<br />
-12n<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 149:Messung an Zelle 16 mit Transistor 5c. Zelle ist abgestorben. Keine<br />
Mittelwertbildung (Bild 515 in iso-Datenaufzeichnung).
Strommodulation Transistor [A]<br />
Stimulus [A]<br />
intracelluläre Spannung [mV]<br />
1n<br />
0<br />
-1n<br />
20<br />
10<br />
0<br />
-10<br />
<strong>Anhang</strong> 161<br />
Ergebnisse aus Messungen mit EOS-Transistoren an Herzmuskelzellen<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
-20<br />
10 20 30 40 50 60 70<br />
2n<br />
0<br />
-2n<br />
-4n<br />
-6n<br />
-8n<br />
-10n<br />
time [ms]<br />
-12n<br />
10 20 30 40 50 60 70<br />
time [ms]<br />
Abb. 150:Messung an Zelle 16 mit Transistor 5c. Zelle ist abgestorben. Mittelwert<br />
über 10 Messungen (Bilder 515-524 in iso-Datenaufzeichnung).
Literaturverzeichnis<br />
Literatur 162<br />
[1] Schmidt, Thews, Physiologie des Menschen, Springer-Lehrbuch, 24. Auflage<br />
[2] Gauer, Kramer, Jung, Allgemeine Neurophysiologie, Band 10, Urban &<br />
Schwarzenberg, München, (1980)<br />
[3] Orgelmeister, Skript Medizinelektronik, <strong>TU</strong>-<strong>Berlin</strong><br />
[4] Instruction Manual Lock-In Amplifier Model 5208, Princeton Applied<br />
Research Corporation, (1988)<br />
[5] M. Reisch, Elektronische Bauelemente, Funktion, Grundschaltungen, Modellierung<br />
mit SPICE, Springer-Verlag, (1998)<br />
[6] A.L. Hodgkin and A.F. Huxley, A qualitative description of membrane current<br />
and its application to conduction and existation in nerve, J.Physiol. 117, 500-<br />
544, (1952)<br />
[7] P. Fromherz, A.Offenhäuser, T.Vetter, J.Weis, A neuron-silicon-junction: A<br />
retzius cell of the leech on an insulated-gate field-effect-transistor, Science<br />
252, 1290-1293, (1991)<br />
[8] R. Weis, Neuron-Transistor-Kopplung: Bestimmung der elektrischen Übertragungseigenschaften,<br />
Universität Ulm Biophysik, (1994)<br />
[9] A. Stett, Extrazelluläre kapazitive Stimulation und Detektion elektrischer<br />
Signale individueller Nervenzellen mit planaren Silizium-Mikrostrukturen,<br />
Dissertation, MPI für Biochemie, Martinsried/München, (1995)<br />
[10] P. Fromherz, A. Strett, Silicon-neuron junction: capazitive stimulation of an<br />
individual neuron on a silicon chip, Phys. Rev. Lett. 75, 1670, (1995)<br />
[11] R. Weis, B.Müller, P.Fromherz, Neuron adhesion on a silicon chip probed by<br />
an array of field-effect transistors, Phys. Rev. Lett. 76, 327, (1996)<br />
[12] R. Weis, P.Fromherz, Frequency dependent signal transfer in neuron transistors,<br />
Phys. Rev. E 55, 877, (1997)<br />
[13] A. Stett, B.Müller, P.Fromherz, Two-way silicon-neuron interface by electrical<br />
induction, Phys. Rev. E 55, 1779, (1997)<br />
[14] P. Fromherz, Self-gating of ion channels in cell adhension, Phys. Rev. Lett.<br />
78, 4131, (1997)<br />
[15] S. Vassanelli, P.Fromherz, Neurons from rat brain coupled to transistors,<br />
Appl. Phys. A 65, 85-88, (1997)<br />
[16] M. Jenkner, P.Fromherz, Bistability of membran conductance in cell adhesion<br />
observed in a neuron transistor, Phya. Rev. Lett. 79, 4705, (1997)<br />
[17] R. Schätzthauer, P.Fromherz, Neuron-silicon junction with voltage-gated<br />
ionic currents, Eur. J. Neurosci. 10, 1956, (1998)<br />
[18] S. Vassanelli, P.Fromherz, Transistor records of excitable neurons from rat<br />
brain, Appl. Phys. A 66, 459-463, (1998)<br />
[19] D. Braun, P.Fromherz, Fluorescence interference-contrast microscopy of cell<br />
adhesion on oxidized silicon, Appl. Phys. A 65, 341-348, (1997)<br />
[20] D. Braun, P. Fromherz, Fluorescence interferometry of neuronal cell<br />
adhesion on microstructured silicon, Phys. Rev. Lett. 81, 5241, (1998)<br />
[21] P. Fromherz, Extracellular recording with transistors and the distribution of<br />
ionic conductances in a cell membrane, Eur. Biophys. J. 28, 254-258, (1999)<br />
[22] W.G. Regehr, A long term in vitro silicon-based microelectrode-neuron<br />
connection, IEEE Trans Biomed Eng 35, 1023-1032, (1988)
Literatur 163<br />
[23] J. Pine, Recording action potentials from cultured neurons with extracellular<br />
microcircuit electrodes, J. Neurosci. Methodes 2, 19-31, (1980)<br />
[24] G.W. Gross, Transparent indium-tin oxide electrode patterns for extracellular,<br />
multisite recording in neuronal cultures, J. Neurosci. Methodes 15, 243-<br />
252, (1985)<br />
[25] W.G. Regehr, Sealing cultured invertebrate neurons to embedded dish<br />
electrodes facilitates long-term stimulation and recording, J. Neurosci.<br />
Methodes 30, 91-106, (1989)<br />
[26] R.J.A. Wilson, Simultabeous multisite recording and stimulation of signale<br />
isolated leech neurons using planar extracellular electrode arrays, J.<br />
Neurosci. Methods 53, 101-110, (1994)<br />
[27] C.D.Motchenbacher, J.A.Connelly, Low-noise electronic system design,<br />
John-Wiley & Sons, Inc., (1993)<br />
[28] B.Sakmann, E.Neher, Singel-Channel Recording, Plenum Press, (1983)<br />
[29] T.Kind, Dissertation, in Arbeit, <strong>TU</strong>-<strong>Berlin</strong><br />
[30] D. Kim, Diplomarbeit, in Arbeit, <strong>TU</strong>-<strong>Berlin</strong><br />
[31] H.G. Wagemann, W. Gerlach, Physik und Technologie der Halbleiter-Bauelemente,<br />
Skriptum, <strong>TU</strong>-<strong>Berlin</strong>, (1992)<br />
[32] A.S. Grove, Physics and technology of semiconductor devices, John Wiley<br />
and Sons, Inc., (1967)
Danksagung<br />
Danksagung 164<br />
Zunächst möchte ich Herrn Prof. Dr. -Ing. H.G. Wagemann dafür danken, daß ich<br />
in seinem Institut meine Diplomarbeit über ein sehr interessantes Thema an der<br />
Schnittstelle zwischen <strong>Mikroelektronik</strong>, Halbleiterphysik und Physiologie anfertigen<br />
konnte. Es hat mir die Möglichkeit gegeben, mich mit Problemstellungen der unterschiedlichen<br />
Fachrichtungen auseinanderzusetzen und mich an der Grundlagenforschung<br />
in einem sehr zukunftsträchtigen Bereich zu beteiligen. Für die intensive<br />
Betreuung meiner Arbeit danke ich Herrn B. Müller. Er war für mich stets der<br />
richtige Ansprechpartner, um über Fortschritte und Rückschläge zu diskutieren.<br />
Herrn B. Müller verdanke ich die technischen Voraussetzungen für diese Arbeit.<br />
Besonderer Dank gilt meinem Betreuer R. Arnold. Seiner großen Geduld und<br />
Bereitschaft auf meine Fragen und Probleme einzugehen und seiner Unterstützung<br />
bei den Messungen verdanke ich den Erfolg meiner Arbeit. In der Zusammenarbeit<br />
mit R. Arnold sind in einem besonders offenen und ungezwungenen Arbeitsklima<br />
viele neuen Ideen entstanden, die den Durchbruch bei den Messungen ermöglicht<br />
haben. Ich erinnere mich sehr gerne an die angeregten Diskussionen und an die<br />
langen Nächte bei den Messungen im Labor.<br />
T. Kind verdanke ich den Aufbau der Infrastruktur, den Entwurf der verwendeten<br />
Transistoren und die intensive Einführung in die Präparation und das Patchen von<br />
Herzmuskelzellen. Ich möchte mich auch bei D. Kim für die Hinweise und Tips zum<br />
Thema Rauschen und für die Durchsicht dieses Teils in meiner Arbeit bedanken.<br />
Die Durchführung dieser Diplomarbeit war nur in einer engen Zusammenarbeit mit<br />
der Medizinischen Fakultät der Humboldt Universität zu <strong>Berlin</strong> (Charité), Institut für<br />
Physiologie, Abteilung Neurophysiologie unter der Leitung von Prof. Dr. med. U.<br />
Heinemann möglich. Für die gute Zusammenarbeit möchte ich mich bedanken.<br />
Mein besonderer Dank gilt dabei besonders Frau C. Gebhardt und Herrn Dr. med.<br />
Günther und Frau S. Latta, die mich mit vielen Ratschlägen zur Präparation und<br />
zum Patchen von Herzmuskelzellen unterstützt haben.<br />
T. Meyer-Thorke danke ich für die Durchsicht der Arbeit auf Rechtschreibung und<br />
allen anderen im Institut für Mikroelekronik und Festkörperelektronik an der Technischen<br />
Universität <strong>Berlin</strong> danke ich für die gute Zusammenarbeit.













