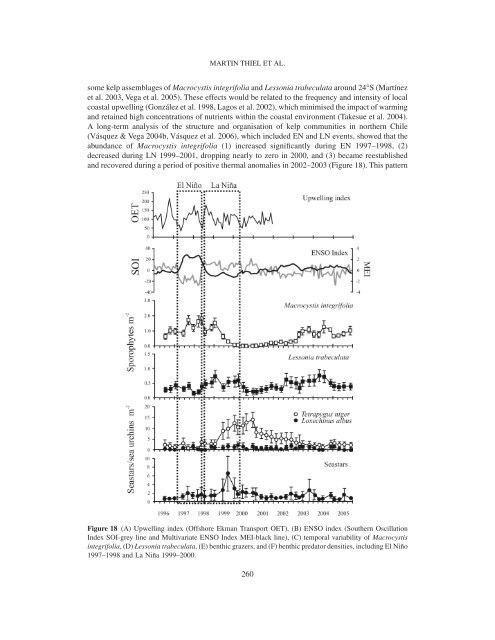
MARTIN THIEL ET AL.some kelp assemblages <strong>of</strong> Macrocystis integrifolia <strong>and</strong> Lessonia trabeculata around 24°S (Martínezet al. 2003, Vega et al. 2005). These effects would be related to <strong>the</strong> frequency <strong>and</strong> intensity <strong>of</strong> localcoastal upwelling (González et al. 1998, Lagos et al. 2002), which minimised <strong>the</strong> impact <strong>of</strong> warming<strong>and</strong> retained high concentrations <strong>of</strong> nutrients within <strong>the</strong> coastal environment (Takesue et al. 2004).A long-term analysis <strong>of</strong> <strong>the</strong> structure <strong>and</strong> organisation <strong>of</strong> kelp communities in nor<strong>the</strong>rn Chile(Vásquez & Vega 2004b, Vásquez et al. 2006), which included EN <strong>and</strong> LN events, showed that <strong>the</strong>abundance <strong>of</strong> Macrocystis integrifolia (1) increased significantly during EN 1997–1998, (2)decreased during LN 1999–2001, dropping nearly to zero in 2000, <strong>and</strong> (3) became reestablished<strong>and</strong> recovered during a period <strong>of</strong> positive <strong>the</strong>rmal anomalies in 2002–2003 (Figure 18). This patternFigure 18 (A) Upwelling index (Offshore Ekman Transport OET), (B) ENSO index (Sou<strong>the</strong>rn OscillationIndex SOI-grey line <strong>and</strong> Multivariate ENSO Index MEI-black line), (C) temporal variability <strong>of</strong> Macrocystisintegrifolia, (D) Lessonia trabeculata, (E) benthic grazers, <strong>and</strong> (F) benthic predator densities, including El Niño1997–1998 <strong>and</strong> La Niña 1999–2000.260
THE HUMBOLDT CURRENT SYSTEM OF NORTHERN AND CENTRAL CHILEwas different from that recorded on <strong>the</strong> California coast, where <strong>the</strong> rapid recovery <strong>of</strong> M. pyriferafollowing EN 1997–1998 was favoured by <strong>the</strong> establishment <strong>of</strong> a cold period (1998–2000) <strong>and</strong> <strong>the</strong>survival <strong>of</strong> sporophytes in deep environments (Ladah et al. 1999; Edwards 2004). In nor<strong>the</strong>rn Chile,<strong>the</strong> recolonisation rate <strong>of</strong> kelp assemblages occurred comparatively slowly (Martínez et al. 2003;see also Population connectivity, p. 252ff. <strong>and</strong> Biogeography, p. 255ff.), even though cold conditionsprevailing during 1998–2000 enhanced <strong>the</strong> upwelling effect. In this regard, <strong>the</strong> slow recovery <strong>of</strong>Lessonia nigrescens after EN 1982–1983 (Castilla & Camus 1992) appeared more related to bioticconstraints: recruitment was strongly reduced by a combination <strong>of</strong> postsettlement grazing <strong>and</strong>inhibition by encrusting coralline algae, while erect coralline algae played a key role as facilitators,allowing <strong>the</strong> kelp some escape from grazers <strong>and</strong> space competitors (Camus 1994a).On <strong>the</strong> o<strong>the</strong>r h<strong>and</strong>, <strong>the</strong> decreased abundance <strong>of</strong> Macrocystis integrifolia was caused by asignificant reduction in <strong>the</strong> adult plant population <strong>and</strong> <strong>the</strong> lack <strong>of</strong> recruitment <strong>of</strong> juvenile sporophytes(Figure 18). Thus, <strong>the</strong> disappearance <strong>of</strong> <strong>the</strong> M. integrifolia population occurred 2 yr after EN1997–1998 <strong>and</strong> was inversely correlated with a population increase <strong>of</strong> <strong>the</strong> sea urchin Tetrapygusniger (Figure 18). In contrast, information from o<strong>the</strong>r areas <strong>of</strong> <strong>the</strong> sou<strong>the</strong>astern Pacific during EN1997–1998 showed that superficial warming decreased <strong>the</strong> abundance <strong>of</strong> kelp on shallow bottoms,inducing migrations <strong>of</strong> grazers to deeper zones (Fernández et al. 1999, Godoy 2000, Lleellish et al.2001). In nor<strong>the</strong>rn Chile, during EN 1997–1998 <strong>and</strong> LN 1998–2000, different events favoured <strong>the</strong>increase <strong>of</strong> sea urchin populations during <strong>the</strong> cold phase, including (1) induction <strong>of</strong> mass spawningdue to increases in SST <strong>and</strong> persistence <strong>of</strong> upwelling events, (2) reduction in density <strong>of</strong> adultseastars, <strong>and</strong> (3) changes in <strong>the</strong> feeding behaviour <strong>of</strong> <strong>the</strong> seastar Heliaster helianthus, one <strong>of</strong> <strong>the</strong>most important benthic predators on Chilean <strong>and</strong> Peruvian coasts (Tokeshi & Romero 1995b,Vásquez et al. 2006) (Figure 18). Thus, <strong>the</strong> long-term study <strong>of</strong> subtidal communities suggests thatdifferent bottom-up <strong>and</strong> top-down factors might control eco<strong>system</strong> changes in nor<strong>the</strong>rn Chile,including (1) <strong>the</strong> intensity <strong>and</strong> frequency <strong>of</strong> upwelling, which may buffer <strong>the</strong> positive <strong>the</strong>rmalanomalies <strong>of</strong> SST <strong>and</strong> maintain high nutrient levels, favouring kelp persistence during EN events;(2) site-dependent oceanographic conditions, which may generate optimal conditions for spawning,larval development, <strong>and</strong> recruitment <strong>of</strong> echinoderms during <strong>and</strong>/or after EN events; (3) an overallabundance increase <strong>of</strong> carnivores which is correlated with an abundance decline <strong>of</strong> <strong>the</strong> mostconspicuous grazers; (4) population dynamics <strong>of</strong> adult seastars <strong>and</strong> sea urchins which may becomedecoupled during EN events; (5) species-specific population dynamics <strong>of</strong> some predator species(e.g., Luidia magellanica), <strong>and</strong> changes in dietary composition in o<strong>the</strong>rs (e.g., H. helianthus), whichmay promote population increase <strong>of</strong> its prey, <strong>the</strong> urchin Tetrapygus niger, during EN events; <strong>and</strong>(6) changes in abundance <strong>of</strong> T. niger, which might be a key factor controlling <strong>the</strong> development <strong>of</strong>two alternate states: environments dominated by kelp beds versus barren ground areas.In a wider context involving both subtidal <strong>and</strong> intertidal environments, EN impacts can besummarised as a large-scale bottom-up effect influencing various (<strong>and</strong> as yet difficult-to-predict)levels <strong>of</strong> marine food webs. However, this is just <strong>the</strong> initial path for most impacts, <strong>and</strong> top-downeffects should not be neglected (e.g., see Nielsen & Navarrete 2004). Future research on EN impactscould consider at least five aspects related to <strong>the</strong> variability <strong>of</strong> biological effects, which may serveas guidelines or study framework: (1) <strong>the</strong> southward intensity attenuation <strong>of</strong> EN signals producesa latitudinal impact gradient, with reduced effects toward higher latitudes (e.g., Castilla & Camus1992, Martínez et al. 2003); (2) in <strong>the</strong> spatiotemporal context, many effects are episodic <strong>and</strong>/orlocal (e.g., abundance variability), <strong>and</strong> some o<strong>the</strong>rs may propagate <strong>the</strong>ir effects to larger spatialscales (e.g., distribution changes, local extinctions), being highly persistent over time (e.g., seeCamus et al. 1994); (3) on a taxonomic basis, some taxa are re<strong>current</strong>ly affected (e.g., kelps), o<strong>the</strong>rsexhibit no significant impacts (e.g., chlorophytes), <strong>and</strong> some taxa can be more affected in <strong>the</strong>irreproduction while o<strong>the</strong>rs in <strong>the</strong>ir recruitment (e.g., Camus 1994a, Navarrete et al. 2005, Vásquezet al. 2006); (4) <strong>the</strong> genetic <strong>and</strong> evolutionary consequences <strong>of</strong> re<strong>current</strong> phenomena such as mass261