

Chapter 5 Genetic Analysis of Apomixis - cimmyt
Chapter 5 Genetic Analysis of Apomixis - cimmyt
Chapter 5 Genetic Analysis of Apomixis - cimmyt

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
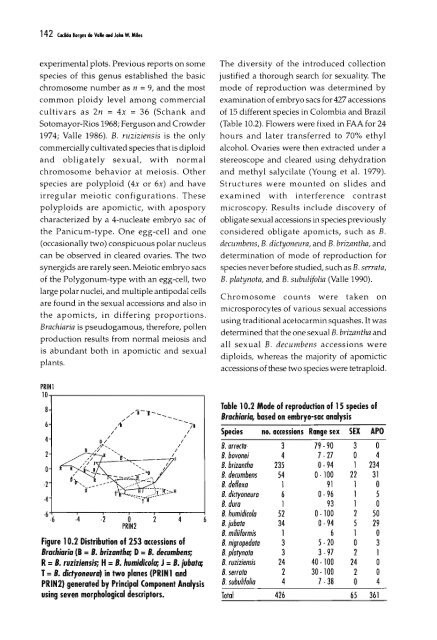
142 C«ida Jorge. do Va" aod Jah W. Milesexperimental plots. Previous reports on somespecies <strong>of</strong> this genus established the basicchromosome number as n = 9, and the mostcommon ploidy level among commercialcultivars as 2n = 4x = 36 (Schank andSotomayor-Rios 1968; Ferguson and Crowder1974; Valle 1986). B. ruziziensis is the onlycommercially cultivated species that is diploidand obligately sexual, with normalchromosome behavior at meiosis. Otherspecies are polyploid (4x or 6x) and haveirregular meiotic configurations. Thesepolyploids are apomictic, with aposporycharacterized by a 4-nucleate embryo sac <strong>of</strong>the Panicum-type. One egg-cell and one(occasionally two) conspicuous polar nucleuscan be observed in cleared ovaries. The twosynergids are rarely seen. Meiotic embryo sacs<strong>of</strong> the Polygonum-type with an egg-cell, twolarge polar nuclei, and multiple antipodal cellsare found in the sexual accessions and also inthe apomicts, in differing proportions.Brachiaria is pseudogamous, therefore, pollenproduction results from normal meiosis andis abundant both in apomictic and sexualplants.PRINI10.,.----------------,·2·4.6.6·4 ·2 oPRIN2Figure 10.2 Distribution <strong>of</strong> 253 accessions <strong>of</strong>Brachiaria (8 =B. brizantha; D=B. decumbens;R= B. ruziziensis; H= B. humidico/a; J = B. jubatc;T= B. didyoneura) in two planes (PRIN 1 andPRIN2) generated by Principal Component <strong>Analysis</strong>using seven morphological descriptors.The diversity <strong>of</strong> the introduced collectionjustified a thorough search for sexuality. Themode <strong>of</strong> reproduction was determined byexamination <strong>of</strong> embryo sacs for 427 accessions<strong>of</strong> 15 different species in Colombia and Brazil(Table 10.2). Flowers were fixed in FAA for 24hours and later transferred to 70% ethylalcohol. Ovaries were then extracted under astereoscope and cleared using dehydrationand methyl salycilate (Young et al. 1979).Structures were mounted on slides andexamined with interference contrastmicroscopy. Results include discovery <strong>of</strong>obligate sexual accessions in species previouslyconsidered obligate apomicts, such as B.decumbens, B. dictyaneura, and B. brizantha, anddetermination <strong>of</strong> mode <strong>of</strong> reproduction forspecies never before studied, such as B. serra/a,B. platynota, and B. subulifolia (Valle 1990).Chromosome counts were taken onmicrosporocytes <strong>of</strong> various sexual accessionsusing traditional acetocarmin squashes. It wasdetermined that the one sexual B. brizantha andall sexual B. decumbens accessions werediplOids, whereas the majority <strong>of</strong> apomicticaccessions <strong>of</strong> these two species were tetraploid.Table 10.2 Mode <strong>of</strong> reproduction <strong>of</strong> 15 species <strong>of</strong>Brachiaria, based on embryo-sac analysisSpedes no. accessions Range sex SEX APOB. affecta·· 3 79 -90 3 oB. bovonei 4 7·27 o 4B. brizantha 235 0- 94 1 234B. decumbens 54 0·100 22 31B. deflexa 1 91 1 oB. dictyoneura 6 0·96 1 5B. dura 1 93 1 oB. humidicola 52 0·100 2 50B. iubata 34 0·94 5 29B. miliiFormis 1 6 1 oB. nigropedata 3 5- 20 o 3B. platynoto 3 3-97 2 1B. ruziziensis 24 40-100 24 oB. serrato 2 30 100 2 oB. subuliFolia 4 7-38 o 4Total 426 65 361













