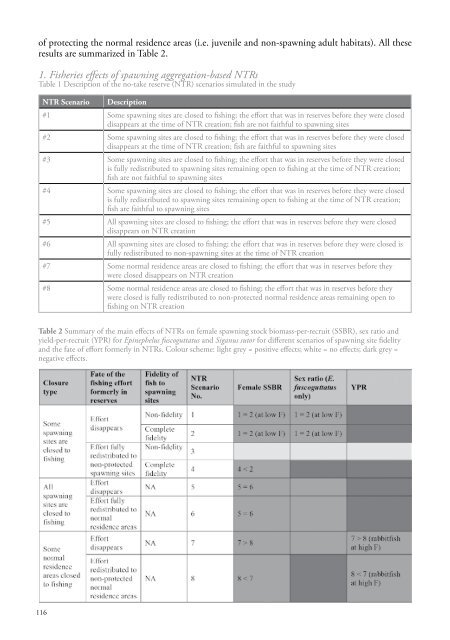
of protecting the normal residence areas (i.e. juvenile and non-<strong>spawning</strong> adult habitats). All theseresults are summarized in Table 2.1. Fisheries effects of <strong>spawning</strong> aggregation-based NTRsTable 1 Description of the no-take reserve (NTR) scenarios simulated in the studyNTR Scenario Description#1 Some <strong>spawning</strong> sites are closed to fishing; the effort that was in reserves before they were closeddisappears at the time of NTR creation; fish are not faithful to <strong>spawning</strong> sites#2 Some <strong>spawning</strong> sites are closed to fishing; the effort that was in reserves before they were closeddisappears at the time of NTR creation; fish are faithful to <strong>spawning</strong> sites#3 Some <strong>spawning</strong> sites are closed to fishing; the effort that was in reserves before they were closedis fully redistributed to <strong>spawning</strong> sites remaining open to fishing at the time of NTR creation;fish are not faithful to <strong>spawning</strong> sites#4 Some <strong>spawning</strong> sites are closed to fishing; the effort that was in reserves before they were closedis fully redistributed to <strong>spawning</strong> sites remaining open to fishing at the time of NTR creation;fish are faithful to <strong>spawning</strong> sites#5 All <strong>spawning</strong> sites are closed to fishing; the effort that was in reserves before they were closeddisappears on NTR creation#6 All <strong>spawning</strong> sites are closed to fishing; the effort that was in reserves before they were closed isfully redistributed to non-<strong>spawning</strong> sites at the time of NTR creation#7 Some normal residence areas are closed to fishing; the effort that was in reserves before theywere closed disappears on NTR creation#8 Some normal residence areas are closed to fishing; the effort that was in reserves before theywere closed is fully redistributed to non-protected normal residence areas remaining open tofishing on NTR creationTable 2 Summary of the main effects of NTRs on female <strong>spawning</strong> stock biomass-per-recruit (SSBR), sex ratio andyield-per-recruit (YPR) for Epinephelus fuscoguttatus and Siganus sutor for different scenarios of <strong>spawning</strong> site fidelityand the fate of effort formerly in NTRs. Colour scheme: light grey = positive effects; white = no effects; dark grey =negative effects.116
• Patterns of YPR in presence of NTRs were qualitatively similar for grouper and rabbitfish.• For all scenarios examined, encompassing both the protection of a fraction or all<strong>spawning</strong> aggregations, there are no benefits for YPR and even negative effects undermost scenarios.• Only if effort is much higher than is currently applied in the trap fishery for rabbitfish,equating to an overexploited stock, can pre-reserve levels of YPR be maintained byprotecting a fraction of <strong>spawning</strong> aggregations.• For the grouper population, YPR decreases with the fraction of <strong>spawning</strong> areas protectedand, the higher the value of effort, the more YPR decreases with the fraction of <strong>spawning</strong>sites in reserves.• The greatest declinesin YPR occur when all <strong>spawning</strong> aggregations are protected, sinceglobal fishing mortality is considerably reduced in this situation, both when effortdisappears and when effort is redistributed.• The negative effects on YPR are more pronounced for grouper than rabbitfish, sincecatchability increases to a larger extent with grouper aggregation formation.2. Conservation effects of protecting a fraction of <strong>spawning</strong> aggregations using NTRs• Patterns of female SSBR in the presence of NTRs were qualitatively similar for grouperand rabbitfish.• Unsurprisingly, the greatest conservation benefits (i.e. increases in SSBR and decreasesin sex ratio) were obtained when effort previously expended on <strong>spawning</strong> aggregationssimply disappeared when the NTRs were established. Interestingly, female SSBR and sexratio were relatively unchanged between the site fidelity and non-site fidelity scenarioswhen effort disappears.• However, at higher levels of annual effort than those observed in the fisheries, SSBR andsex ratio would benefit more from the site fidelity than the non-site fidelity situation, asthe fraction of protected <strong>spawning</strong> sites increases.• By contrast, site fidelity has important consequences for conservation benefits if effortat <strong>spawning</strong> sites is redistributed when NTRs are created. As we assumed that effortwould be redistributed to the remaining unprotected <strong>spawning</strong> sites if only a fraction areprotected using NTRs, SSBR is invariant with the fraction of <strong>spawning</strong> sites in reservesin the non-site fidelity scenario, as fish would suffer mortality when they attended nonprotected<strong>spawning</strong> aggregations.• Conversely, if fish are faithful to <strong>spawning</strong> sites, conservation benefits are attained oneffort redistribution, but at lower levels than when effort disappears.• However, the effects of effort redistribution on sex ratio for the protogynous grouperpopulation were negative. At high effort levels and with a relatively large fraction of<strong>spawning</strong> sites protected by NTRs, sex ratio at fished <strong>spawning</strong> sites would be moreheavily female-biased than if the same level of effort was applied in the fishery withoutany NTRs.3. Conservation effects of protecting all <strong>spawning</strong> aggregations using NTRs• If all <strong>spawning</strong> aggregations are protected by NTRs, conservation benefits occur anddo not differ much if effort disappears or is redistributed. This is because, under thisassumption, all effort would be directed towards non-<strong>spawning</strong> periods and areas wherecatchability is much lower. Therefore, even the addition of redistributed effort on non<strong>spawning</strong>fish does not affect SSBR to a large degree.• There are greater conservation benefits for groupers than rabbitfish, since in the former,aggregation formation is associated with much larger increases in relative catchability.• The benefits for male groupers are also higher than those for females – as males spendlonger times aggregating, total closure of <strong>spawning</strong> sites increases SSB for males to alarger extent than for females.117
- Page 4:
The designation of geographical ent
- Page 9:
Chapter 1: IntroductionJan Robinson
- Page 12 and 13:
limited, subsistence levels of expl
- Page 14:
NTRs for spawning aggregations usin
- Page 17 and 18:
al. 2003). Verification may include
- Page 19 and 20:
a fraction of spawning sites are pr
- Page 21 and 22:
Chapter 3: Targeted fishing of the
- Page 23 and 24:
verifying spawning aggregations, we
- Page 25 and 26:
ecorded from inshore close to the c
- Page 27 and 28:
(a)(b)Fig. 3. Spatial patterns ofca
- Page 30 and 31:
pooled sizes of the three spawning
- Page 32 and 33:
found S. sutor contributed up to 44
- Page 34 and 35:
2011b). However, observations of fi
- Page 36 and 37:
MethodsTo identify seasonal and lun
- Page 38 and 39:
n=199Females GSI (mean ± SE)2.521.
- Page 40 and 41:
The estimate of size at maturity in
- Page 42 and 43:
This study was designed to verify S
- Page 44 and 45:
were selected. Fish selected for ta
- Page 46 and 47:
The number of traps increased on th
- Page 48 and 49:
Of the 9 tagged fish detected by re
- Page 50 and 51:
Fig. 7. Diel patterns ofdetection f
- Page 52 and 53:
Spawning aggregation site fidelity
- Page 54 and 55:
Chapter 6: Shoemaker spinefoot rabb
- Page 56 and 57:
anterior of the anus and below the
- Page 58 and 59:
A high percentage (80.8%) of depart
- Page 60 and 61:
arrivals and departures at these tw
- Page 62 and 63:
are typically applied for reef fish
- Page 64 and 65:
(a)(b)(c)Chapter 3, Figure 3. Spati
- Page 66 and 67:
(1)(2)(3)(4)(5)(6)Chapter 7, Table
- Page 68 and 69:
Chapter 12, Fig. 1 Fraction of fema
- Page 70 and 71:
Plates 8. Selected photographs from
- Page 72 and 73:
MethodsStudy sitesThe study area wa
- Page 74 and 75: which shelved gently ( ca. 25 o ) t
- Page 76 and 77: Fig. 4. Lunar periodicity in number
- Page 78 and 79: Behaviour and appearanceDescription
- Page 80 and 81: eported aggregations forming betwee
- Page 82 and 83: The sizes of E. fuscoguttatus aggre
- Page 84 and 85: Materials and methodsStudy area and
- Page 86 and 87: TL. All fish tagged were considered
- Page 88 and 89: Lunar timing of arrivals and depart
- Page 90 and 91: Fig. 8. The presence and absence of
- Page 92 and 93: aggregation fishing. This critical
- Page 94 and 95: Chapter 9: Persistence of grouper (
- Page 96 and 97: ResultsBetween 2003 and 2006, the c
- Page 98 and 99: Fig. 2. Mean (± standard error, SE
- Page 100 and 101: A few species (e.g. Epinephelus gut
- Page 102 and 103: (a)(b)Fig. 1. Map of (a) study site
- Page 104 and 105: Fig. 2. Number of E. lanceolatus ob
- Page 106 and 107: was having any impact on the popula
- Page 108 and 109: A spawning aggregation is said to o
- Page 110 and 111: Chapter 11: Evaluation of an indica
- Page 112 and 113: Table 1 Aggregation fisheries asses
- Page 114 and 115: the lists of Jennings et al. (1999)
- Page 116 and 117: with the more vulnerable labrids an
- Page 118 and 119: The remaining serranid populations
- Page 120 and 121: Spawning aggregation behaviour is c
- Page 122 and 123: tiger grouper, Mycteroperca tigris:
- Page 126 and 127: • Since grouper males are afforde
- Page 128 and 129: Fig. 2 Yield-per-recruit normalized
- Page 130 and 131: The approaches identified above are
- Page 132 and 133: during full moon periods. Siganus s
- Page 134 and 135: model, many parameter estimates are
- Page 136 and 137: ReferencesAbunge C (2011) Managing
- Page 138 and 139: Cox DR (1972) Regression models and
- Page 140 and 141: Grüss A, Kaplan DM, Hart DR (2011b
- Page 142 and 143: Kaunda-Arara B, Rose GA (2004a) Eff
- Page 144: Newcomer RT, Taylor DH, Guttman SI
- Page 147 and 148: Sancho G, Petersen CW, Lobel PS (20
- Page 149 and 150: Appendix 1. QuestionnaireMASMA SPAW
- Page 151 and 152: 8. Spawning aggregation knowledgeUs
- Page 153 and 154: Example items KSh Furthest site Clo
- Page 155 and 156: Appendix II. Experimental testing o
- Page 157 and 158: Clove oil concentrationAt a concent
- Page 159 and 160: Appendix III. Application of acoust
- Page 161 and 162: 153