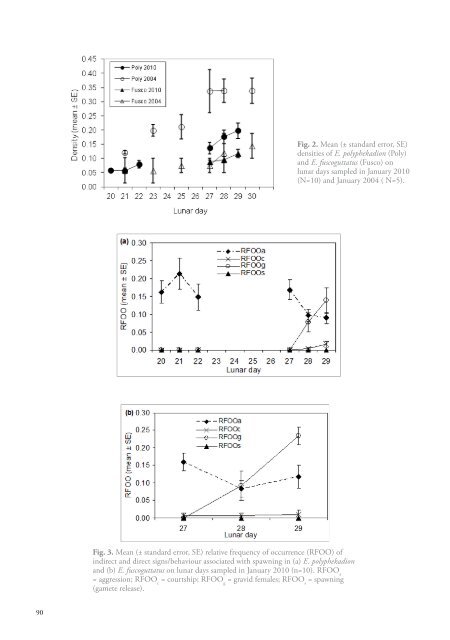
Fig. 2. Mean (± standard error, SE)densities of E. polyphekadion (Poly)and E. fuscoguttatus (Fusco) onlunar days sampled in January 20<strong>10</strong>(N=<strong>10</strong>) and January 2004 ( N=5).Fig. 3. Mean (± standard error, SE) relative frequency of occurrence (RFOO) ofindirect and direct signs/behaviour associated with <strong>spawning</strong> in (a) E. polyphekadionand (b) E. fuscoguttatus on lunar days sampled in January 20<strong>10</strong> (n=<strong>10</strong>). RFOO a= aggression; RFOO c= courtship; RFOO g= gravid females; RFOO s= <strong>spawning</strong>(gamete release).90
with the arrival of gravid females, whereas E. fuscoguttatus aggression remained relatively stable overthe few days this species was surveyed. Courtship was rarely observed but occurrences increased onthe last day of the surveys. Spawning rushes and gamete release were not observed.DiscussionThe most likely cause of the large-scale loss and degradation of <strong>spawning</strong> site habitat was CycloneBondo, which struck the atoll on 22 December 2006 (Chang-Seng 2007). Cyclones are infrequent(decadal-scale) and generally of low intensity in Seychelles’ southern atoll groups (Chang-Seng2007). While reef degradation from sand inundation might be explained by changes in current orwave patterns, or increased rates of carbonate erosion, such processes typically operate on muchlonger time scales (Woodroffe 2003). Further visual evidence for a massive cyclone impact alongthe northern edge of the atoll included recent deposits of coral boulders on the reef crest and loss oraccumulation of emergent sand banks. Moreover, fishers at the atoll report larger waves and moredifficult navigation in the pass since the cyclone, possibly a result of sand accumulation.Four years after the cyclone, large <strong>spawning</strong> aggregations of both species continue to form at thesite. Based on known seasonality and lunar periodicity of aggregations at the study site (Robinsonet al. 2008), <strong>spawning</strong> and dispersal of the December 2006 aggregations are likely to have occurreda few days prior to the impact of the storm. Fish departing the aggregation site were also unlikelyto have suffered direct mortality as a result of the cyclone since large and mobile reef fishes aretypically able to avoid such impacts (Lassig 1983). If immediate lethal effects of the cyclone on<strong>spawning</strong> and migrating adults are considered negligible, the potential for longer-term impactsmay depend on the importance of habitat quality for aggregation formation and <strong>spawning</strong> success.Aggregating E. polyphekadion and E. fuscoguttatus compensated for the considerable habitatdisturbance through a redistribution of <strong>spawning</strong> areas, whereby the core of the aggregationsshifted to the nearest available hard substrate habitat that bordered the reef pass. This suggests thatspecific features (e.g. individual coral heads) may not be critical for aggregation persistence in thesespecies. Nonetheless, both species clearly require reef and a degree of coral structure for <strong>spawning</strong>aggregation formation, since fish redistributed themselves to coral areas rather than returningto the sand-inundated half of the core site. Consequently, <strong>spawning</strong> aggregation persistence isunlikely to be threatened by disturbances as long as reef areas remain available. However, we wereunable to formally compare densities or abundances and it is possible that aggregation sizes mayhave changed following the disturbance. Moreover, the observed habitat changes may have affectedreproductive output if, for example, female selection of male territories is based on specific hardsubstrate attributes, such as level of rugosity.The mechanisms that enable persistence of aggregations at a site are of immediate concern formanagement of aggregation fisheries (Sadovy and Domeier 2005). Although they are not wellunderstood, Warner (1988) identifies the potential role of tradition and social behaviour in <strong>spawning</strong>aggregation persistence, whereby young adult fish learn from older fish to identify established<strong>spawning</strong> sites. Since the aggregating populations at our study site appeared unperturbed by thecyclone, at least in the medium-term, any social behaviour enabling traditional use of the site seemsto have remained intact. Also illustrated by the manipulative experiment of Warner (1988) is thefact that traditional use of a site appears to override specific habitat attributes, which is a patternsupported by our results. While traditional site use was maintained, many individuals arriving atthe site after the cyclone compensated for a reduction in the traditional core (<strong>spawning</strong>) reef area byadopting different parts of the reef for <strong>spawning</strong>. Therefore, the exact location and distribution ofaggregations within a <strong>spawning</strong> site may be dictated by the requirements for suitable habitat ratherthan specific features for territorial, courtship or predator avoidance behaviour, and will depend onhabitat configuration. Location and distribution of aggregations also change with aggregation sizesince they develop from small regions of the core <strong>spawning</strong> area and then occupy increasing area ofhabitat as more fish arrive (JR, pers. observation).91
- Page 4:
The designation of geographical ent
- Page 9:
Chapter 1: IntroductionJan Robinson
- Page 12 and 13:
limited, subsistence levels of expl
- Page 14:
NTRs for spawning aggregations usin
- Page 17 and 18:
al. 2003). Verification may include
- Page 19 and 20:
a fraction of spawning sites are pr
- Page 21 and 22:
Chapter 3: Targeted fishing of the
- Page 23 and 24:
verifying spawning aggregations, we
- Page 25 and 26:
ecorded from inshore close to the c
- Page 27 and 28:
(a)(b)Fig. 3. Spatial patterns ofca
- Page 30 and 31:
pooled sizes of the three spawning
- Page 32 and 33:
found S. sutor contributed up to 44
- Page 34 and 35:
2011b). However, observations of fi
- Page 36 and 37:
MethodsTo identify seasonal and lun
- Page 38 and 39:
n=199Females GSI (mean ± SE)2.521.
- Page 40 and 41:
The estimate of size at maturity in
- Page 42 and 43:
This study was designed to verify S
- Page 44 and 45:
were selected. Fish selected for ta
- Page 46 and 47:
The number of traps increased on th
- Page 48 and 49: Of the 9 tagged fish detected by re
- Page 50 and 51: Fig. 7. Diel patterns ofdetection f
- Page 52 and 53: Spawning aggregation site fidelity
- Page 54 and 55: Chapter 6: Shoemaker spinefoot rabb
- Page 56 and 57: anterior of the anus and below the
- Page 58 and 59: A high percentage (80.8%) of depart
- Page 60 and 61: arrivals and departures at these tw
- Page 62 and 63: are typically applied for reef fish
- Page 64 and 65: (a)(b)(c)Chapter 3, Figure 3. Spati
- Page 66 and 67: (1)(2)(3)(4)(5)(6)Chapter 7, Table
- Page 68 and 69: Chapter 12, Fig. 1 Fraction of fema
- Page 70 and 71: Plates 8. Selected photographs from
- Page 72 and 73: MethodsStudy sitesThe study area wa
- Page 74 and 75: which shelved gently ( ca. 25 o ) t
- Page 76 and 77: Fig. 4. Lunar periodicity in number
- Page 78 and 79: Behaviour and appearanceDescription
- Page 80 and 81: eported aggregations forming betwee
- Page 82 and 83: The sizes of E. fuscoguttatus aggre
- Page 84 and 85: Materials and methodsStudy area and
- Page 86 and 87: TL. All fish tagged were considered
- Page 88 and 89: Lunar timing of arrivals and depart
- Page 90 and 91: Fig. 8. The presence and absence of
- Page 92 and 93: aggregation fishing. This critical
- Page 94 and 95: Chapter 9: Persistence of grouper (
- Page 96 and 97: ResultsBetween 2003 and 2006, the c
- Page 100 and 101: A few species (e.g. Epinephelus gut
- Page 102 and 103: (a)(b)Fig. 1. Map of (a) study site
- Page 104 and 105: Fig. 2. Number of E. lanceolatus ob
- Page 106 and 107: was having any impact on the popula
- Page 108 and 109: A spawning aggregation is said to o
- Page 110 and 111: Chapter 11: Evaluation of an indica
- Page 112 and 113: Table 1 Aggregation fisheries asses
- Page 114 and 115: the lists of Jennings et al. (1999)
- Page 116 and 117: with the more vulnerable labrids an
- Page 118 and 119: The remaining serranid populations
- Page 120 and 121: Spawning aggregation behaviour is c
- Page 122 and 123: tiger grouper, Mycteroperca tigris:
- Page 124 and 125: of protecting the normal residence
- Page 126 and 127: • Since grouper males are afforde
- Page 128 and 129: Fig. 2 Yield-per-recruit normalized
- Page 130 and 131: The approaches identified above are
- Page 132 and 133: during full moon periods. Siganus s
- Page 134 and 135: model, many parameter estimates are
- Page 136 and 137: ReferencesAbunge C (2011) Managing
- Page 138 and 139: Cox DR (1972) Regression models and
- Page 140 and 141: Grüss A, Kaplan DM, Hart DR (2011b
- Page 142 and 143: Kaunda-Arara B, Rose GA (2004a) Eff
- Page 144: Newcomer RT, Taylor DH, Guttman SI
- Page 147 and 148: Sancho G, Petersen CW, Lobel PS (20
- Page 149 and 150:
Appendix 1. QuestionnaireMASMA SPAW
- Page 151 and 152:
8. Spawning aggregation knowledgeUs
- Page 153 and 154:
Example items KSh Furthest site Clo
- Page 155 and 156:
Appendix II. Experimental testing o
- Page 157 and 158:
Clove oil concentrationAt a concent
- Page 159 and 160:
Appendix III. Application of acoust
- Page 161 and 162:
153