Tiergenetische Ressourcen in Deutschland - BMELV
Tiergenetische Ressourcen in Deutschland - BMELV
Tiergenetische Ressourcen in Deutschland - BMELV

Sie wollen auch ein ePaper? Erhöhen Sie die Reichweite Ihrer Titel.
YUMPU macht aus Druck-PDFs automatisch weboptimierte ePaper, die Google liebt.
Annähernd kann davon ausgegangen werden, dass <strong>in</strong><br />
e<strong>in</strong>er zahlenmäßig begrenzten Population der Größe<br />
Ne nach t Generationen Zufalls paarung noch e<strong>in</strong> Anteil<br />
von<br />
73<br />
6 poqo ( 1 – 1<br />
2 Ne<br />
Genorten segregiert, d.h. beide Ausgangsallele noch<br />
enthält.<br />
Was dies für die Erhaltung der genetischen Diversität<br />
<strong>in</strong> Populationen bedeutet, zeigen die Zahlen <strong>in</strong><br />
Tabelle 18.<br />
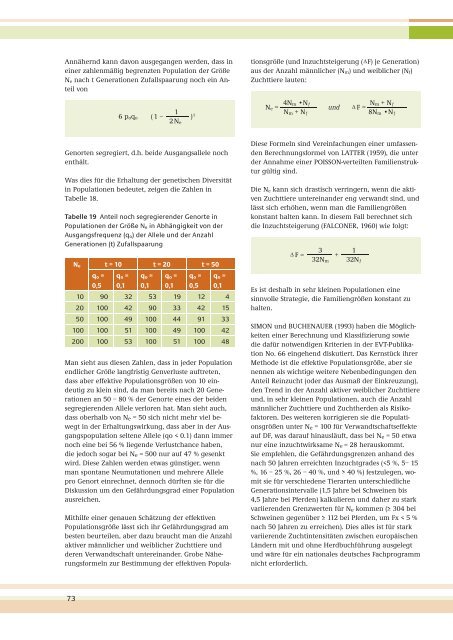
Tabelle 19 Anteil noch segregierender Genorte <strong>in</strong><br />
Populationen der Größe Ne <strong>in</strong> Abhängigkeit von der<br />
Ausgangsfrequenz (qo) der Allele und der Anzahl<br />
Generationen (t) Zufallspaarung<br />
Ne t = 10 t = 20 t = 50<br />
qo =<br />
0,5<br />
qo =<br />
0,1<br />
qo =<br />
0,1<br />
qo =<br />
0,1<br />
qo =<br />
0,5<br />
qo =<br />
0,1<br />
10 90 32 53 19 12 4<br />
20 100 42 90 33 42 15<br />
50 100 49 100 44 91 33<br />
100 100 51 100 49 100 42<br />
200 100 53 100 51 100 48<br />
Man sieht aus diesen Zahlen, dass <strong>in</strong> jeder Population<br />
endlicher Größe langfristig Genverluste auftreten,<br />
dass aber effektive Populationsgrößen von 10 e<strong>in</strong>deutig<br />
zu kle<strong>in</strong> s<strong>in</strong>d, da man bereits nach 20 Generationen<br />
an 50 – 80 % der Genorte e<strong>in</strong>es der beiden<br />
segregierenden Allele verloren hat. Man sieht auch,<br />
dass oberhalb von Ne = 50 sich nicht mehr viel bewegt<br />
<strong>in</strong> der Erhaltungswirkung, dass aber <strong>in</strong> der Ausgangspopulation<br />
seltene Allele (qo < 0.1) dann immer<br />
noch e<strong>in</strong>e bei 56 % liegende Verlustchance haben,<br />
die jedoch sogar bei Ne = 500 nur auf 47 % gesenkt<br />
wird. Diese Zahlen werden etwas günstiger, wenn<br />
man spontane Neumutationen und mehrere Allele<br />
pro Genort e<strong>in</strong>rechnet, dennoch dürften sie für die<br />
Diskussion um den Gefährdungsgrad e<strong>in</strong>er Population<br />
ausreichen.<br />
Mithilfe e<strong>in</strong>er genauen Schätzung der effektiven<br />
Populationsgröße lässt sich ihr Gefährdungsgrad am<br />
besten beurteilen, aber dazu braucht man die Anzahl<br />
aktiver männlicher und weiblicher Zuchttiere und<br />
deren Verwandtschaft untere<strong>in</strong>ander. Grobe Näherungsformeln<br />
zur Bestimmung der effektiven Popula-<br />
) t<br />
tionsgröße (und Inzuchtsteigerung (∆F) je Generation)<br />
aus der Anzahl männlicher (Nm) und weiblicher (Nf)<br />
Zuchttiere lauten:<br />
Ne = 4Nm •Nƒ<br />
Nm + Nƒ<br />
und<br />
∆ F = Nm + Nƒ<br />
8Nm •Nƒ<br />
Diese Formeln s<strong>in</strong>d Vere<strong>in</strong>fachungen e<strong>in</strong>er umfassenden<br />
Berechnungsformel von LATTER (1959), die unter<br />
der Annahme e<strong>in</strong>er POISSON-verteilten Familienstruktur<br />
gültig s<strong>in</strong>d.<br />
Die Ne kann sich drastisch verr<strong>in</strong>gern, wenn die aktiven<br />
Zuchttiere untere<strong>in</strong>ander eng verwandt s<strong>in</strong>d, und<br />
lässt sich erhöhen, wenn man die Familiengrößen<br />
konstant halten kann. In diesem Fall berechnet sich<br />
die Inzuchtsteigerung (FALCONER, 1960) wie folgt:<br />
∆ F =<br />
3<br />
32Nm<br />
+<br />
1<br />
32Nƒ<br />
Es ist deshalb <strong>in</strong> sehr kle<strong>in</strong>en Populationen e<strong>in</strong>e<br />
s<strong>in</strong>nvolle Strategie, die Familiengrößen konstant zu<br />
halten.<br />
SIMON und BUCHENAUER (1993) haben die Möglichkeiten<br />
e<strong>in</strong>er Berechnung und Klassifi zierung sowie<br />
die dafür notwendigen Kriterien <strong>in</strong> der EVT-Publikation<br />
No. 66 e<strong>in</strong>gehend diskutiert. Das Kernstück ihrer<br />
Methode ist die effektive Populationsgröße, aber sie<br />
nennen als wichtige weitere Nebenbed<strong>in</strong>gungen den<br />
Anteil Re<strong>in</strong>zucht (oder das Ausmaß der E<strong>in</strong>kreuzung),<br />
den Trend <strong>in</strong> der Anzahl aktiver weiblicher Zuchttiere<br />
und, <strong>in</strong> sehr kle<strong>in</strong>en Populationen, auch die Anzahl<br />
männlicher Zuchttiere und Zuchtherden als Risikofaktoren.<br />
Des weiteren korrigieren sie die Populationsgrößen<br />
unter Ne = 100 für Verwandtschaftseffekte<br />
auf DF, was darauf h<strong>in</strong>ausläuft, dass bei Ne = 50 etwa<br />
nur e<strong>in</strong>e <strong>in</strong>zuchtwirksame Ne = 28 herauskommt.<br />
Sie empfehlen, die Gefährdungsgrenzen anhand des<br />
nach 50 Jahren erreichten Inzuchtgrades ( 40 %) festzulegen, womit<br />
sie für verschiedene Tierarten unterschiedliche<br />
Generations<strong>in</strong>tervalle (1,5 Jahre bei Schwe<strong>in</strong>en bis<br />
4,5 Jahre bei Pferden) kalkulieren und daher zu stark<br />
variierenden Grenzwerten für Ne kommen (≥ 304 bei<br />
Schwe<strong>in</strong>en gegenüber ≥ 112 bei Pferden, um Fx < 5 %<br />
nach 50 Jahren zu erreichen). Dies alles ist für stark<br />
variierende Zucht<strong>in</strong>tensitäten zwischen europäischen<br />
Ländern mit und ohne Herdbuchführung ausgelegt<br />
und wäre für e<strong>in</strong> nationales deutsches Fachprogramm<br />
nicht erforderlich.