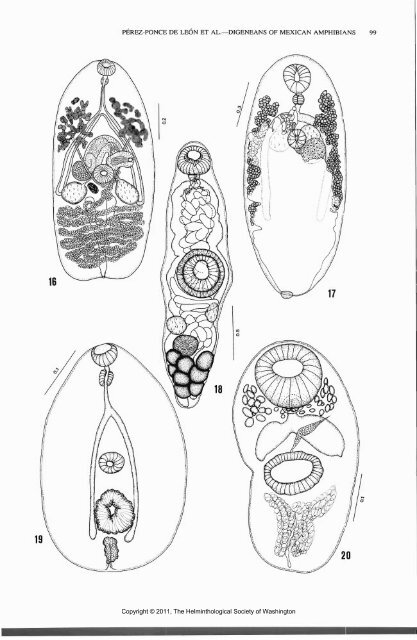
COMPARATIVE PARASITOLOGY, <strong>67</strong>(1), JANUARY <strong>2000</strong> (Leon-Regagnon et al., 1999). Species of Haematoloechus apparently have experienced a diversification in frogs and salamanders in central Mexico, representing the group with the highest species richness (6) in our samples. Glypthelmins, parasitic mainly in frogs of the new world, also shows high species richness. However, the presence at least of 4 species is mainly the result of independent host capture events, either from the neotropical or nearctic zones. The distribution of the nonendemic frogs from the lowlands ranges from Ecuador and Colombia to Veracruz, Mexico (R. vaillanti), to Sonora, northwestern Mexico (L. melanonotus), and Texas (S. baudinii). Their digenean fauna in Veracruz is composed of a combination of neotropical and nearctic species. Only 2 digenean species, Glypthelmins sp. and M. monas, were recovered from L. melanonotus and S. baudinii, respectively. The former probably represents an undescribed species, and M. monas has been reported in numerous host genera in South America and Africa (Prudhoe and Bray, 1982). Seven digenean species were collected from R. vaillanti, and 3 of them show a neotropical distribution: C. rodriguezi in Panama (Caballero, 1955) and G. parva in Brazil (Prudhoe and Bray, 1982), both described from Leptodactylus ocellatus, and G. facioi in Costa Rica (Brenes et al., 1959) and Veracruz, Mexico (Razo-Mendivil et al., 1999), from Rana palmipes Spix, 1824, and R. vaillanti, respectively. The presence of these neotropical digeneans in Los Tuxtlas reflects the geographic distribution of the host genus Leptodactylus and the "Rana palmipes complex" (Frost, 1985; Hillis and De Sa, 1988). One species is endemic, L. (L.) macrocirra, and the 3 remaining species parasitizing R. vaillanti have a nearctic distribution; 2 of these species (G. attenuata and C. americanus) are also present in the endemic frogs of the Transverse Volcanic Axis, and the third species (H. medioplexus) has been recorded in several species of frogs from the United <strong>State</strong>s and Canada, most commonly in members of the "Rana pipiens complex." The 3 species have a low host specificity and have been able to colonize several host groups, thus expanding their distribution range. Apparently, a mixture of neotropical and nearctic species of parasites is taking effect in the lowlands of the Gulf of Mexico, with a series of very interesting phenomena of colonization of new hosts and habitats. Little is known about the amphibian parasite fauna of the tropical lowlands of the Pacific slope of Mexico or the southeastern part of the country. Those areas will undoubtedly be a source of extensive phylogenetic and biogeographic information on parasites and hosts in the future. Contemporary ecological conditions are also important determinants of the parasitic fauna of a host species. Several authors have demonstrated a marked correlation between the relative amount of time spent in association with aquatic habitats and the number of species of platyhelminths hosted by frogs (Brandt, 1936; Prokopic and Krivanec, 1975; Brooks, 1976, 1984; Guillen, 1992). Our data clearly demonstrate that frogs and salamanders harbor the richer digenean fauna compared with the less water-dependent hylids or toads, where digeneans were almost or absolutely absent (the small sample size in leptodactylids precludes any discussion about their helminth fauna). Within frogs and salamanders, diet is the factor that most determines the richness of the digenean communities. Frogs become infected when they prey upon insects or copepods (which is the case in species of Haematoloechus and Halipegus, respectively), when they swallow their own skin bearing encysted metacercariae during ecdysis, or when they feed upon infected tadpoles (species of Catadiscus, Cephalogonimus, Glypthelmins, Gorgoderina, and Megalodiscus) (Yamaguti, 1975; Prudhoe and Bray, 1982). Salamanders of the genus Ambystoma Tschudi, 1838, hosted fewer digenean species than frogs. Garcia-Altamirano et al. (1993) reported that A. dumerilii in Lake Patzcuaro feeds mainly on crayfish and fish. As evidenced by the presence of C. americanus, G. attenuata, and Haematoloechus spp. in salamanders of our samples, it is possible that they oc- Figures 16-20. Ventral views. 16. Loxogenes (I^angeronia) macrocirra (Caballero y Bravo, 1949) Yamaguti, 1971. 17. Mesocoelium monas (Rudolphi, 1819) Teixeira de Freitas, 1958. 18. Halipegus occidualis Stafford, 1905. 19. Fibricola sp. (inetacercaria). 20. Ochetosoma sp. (metacercaria). Scales in millimeters. Copyright © 2011, The Helminthological Society of Washington
PEREZ-PONCE DE LEON ET AL.—DIGENEANS OF MEXICAN AMPHIBIANS 99 Copyright © 2011, The Helminthological Society of Washington 20
- Page 1 and 2:
January 2000 Number 1 Comparative P
- Page 3 and 4:
Comp. Parasitol. 67(1). 2000 pp. 1-
- Page 5 and 6:
children. This means, to most of th
- Page 7 and 8:
liese, 1995; Marcogliese and Cone,
- Page 9 and 10:
ural and human alterations of ecosy
- Page 11 and 12:
ternationally and locally; (2) be i
- Page 13 and 14:
justify the inclusion of parasites
- Page 15 and 16:
phy to understand faunal structure
- Page 17 and 18:
Eucestoda) coincided with the diver
- Page 19 and 20:
(Hoberg et al., 2000). These studie
- Page 21 and 22:
serve biodiversity effectively if e
- Page 23 and 24:
Clayton, D. H., and B. A. Walther.
- Page 25 and 26:
ham. 1996. Combining data in phylog
- Page 27 and 28:
schistosomes. Journal of Parasitolo
- Page 29 and 30:
MARCOGLIESE ET AL.—DIPLOSTOMUM SP
- Page 31 and 32:
MARCOGLIESE HT f^L.—DIPLOSTOMUM S
- Page 33 and 34:
MARCOGLIESE ET AL.—DIPLOSTOMUM SP
- Page 35 and 36:
(Latreille, 1804) Brandt and Ratzeb
- Page 37 and 38:
COADY AND N]CKOL—PLAGIORHYNCHUS C
- Page 39 and 40:
COADY AND NICKOL—PLAGIORHYNCHUS C
- Page 41 and 42:
Threlfall, W. 1965. Helminth parasi
- Page 43 and 44:
strictive traits that were basicall
- Page 45 and 46:
Cement gland usually small with few
- Page 47 and 48:
AMIN ET AL.—REVISION OF THE GENUS
- Page 49 and 50: AMIN ET AL.—REVISION OF THE GENUS
- Page 51 and 52: 17. Trunk spines appearing continuo
- Page 53 and 54: Comp. Parasitol. 67(1), 2000 pp. 51
- Page 55 and 56: SMALES—POPOVASTRONGYLUS FROM MARS
- Page 57 and 58: tion in a circular buccal capsule.
- Page 59 and 60: strongylus pluteus differs from P.
- Page 61 and 62: garoo from Kangaroo Island (Smales
- Page 63 and 64: BURSEY AND GOLDBERG~-/l/VG7aSTOAM O
- Page 65 and 66: Table 2. Parasite list for Onychoda
- Page 67 and 68: phibian helminths in Japan. VI. Pse
- Page 69 and 70: quadrant and 70° in male, 75° in
- Page 71 and 72: increase in the number of cuticular
- Page 73 and 74: Comp. Parasitol. 67(1), 2000 pp. 71
- Page 75 and 76: Figures 1-3. Pseudoterranova decipi
- Page 77 and 78: AMIN ET M^.—PSEUDOTERRANOVA IN MA
- Page 79 and 80: the gill baskets of some hosts were
- Page 81 and 82: KRITSKY ET AL.—DACTYLOGYRIDS FROM
- Page 83 and 84: er, and Boeger, 1986, and Amphoclei
- Page 85 and 86: 1974); P. laticeps Eigenmann, Lagun
- Page 87 and 88: Comp. Parasitol. 67(1), 2000 pp. 85
- Page 89 and 90: MENDOZA-FRANCO ET SCIADICLEITHRUM F
- Page 91 and 92: MENDOZA-FRANCO ET AL.—SCIADICLEIT
- Page 93 and 94: MENDOZA-FRANCO ET M^.—SCIADICLEIT
- Page 95 and 96: PEREZ-PONCE DE LEON ET AL.—DIGENE
- Page 97 and 98: PEREZ-PONCE DE LEON ET AL.—DIGENE
- Page 99: PEREZ-PONCE DE LEON ET AL.—DIGENE
- Page 103 and 104: Table 2. Continued. Locality* (CNHE
- Page 105 and 106: Table 2. Continued. Locality (CNHE
- Page 107 and 108: casionally prey on tadpoles and ins
- Page 109 and 110: Comp. Parasitol. 67(1), 2000 pp. 10
- Page 111 and 112: Comp. Parasitol. 67(1), 2000 pp. 10
- Page 113 and 114: Table 1. Extended. Ctenophorus reti
- Page 115 and 116: Table 2. Extended. 3 '£ Oswaldofil
- Page 117 and 118: Mediorhynchus orientalis Belopol'sk
- Page 119 and 120: 4-5 spines each: 25.0-35.0 (31.4).
- Page 121 and 122: Table 1. Previous helminth records
- Page 123 and 124: hyrae from a collection of lizards.
- Page 125 and 126: WEST ET AL.—RESEARCH NOTES 123 Ta
- Page 127 and 128: 1987, and from these only a sample
- Page 129 and 130: snakes are part of their diet. The
- Page 131 and 132: Comp. Parasitol. 67(1), 2000 pp. 12
- Page 133 and 134: Table 2. Published records of helmi
- Page 135 and 136: , , B. K. Sullivan, and Q. A. Truon
- Page 137 and 138: were larvae. Still, we have conclud
- Page 139 and 140: and their habitats. These low paras
- Page 141 and 142: e eligible for election to office.
- Page 143 and 144: New business. Presentation of notes
- Page 145 and 146: Name: MEMBERSHIP APPLICATION 143 AP
- Page 147 and 148: *Edna M. Buhrer *Mildred A. Doss *A