

Role of the ubiquitin-like modifier FAT10 in protein degradation and ...
Role of the ubiquitin-like modifier FAT10 in protein degradation and ...
Role of the ubiquitin-like modifier FAT10 in protein degradation and ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
Chapter 3<br />
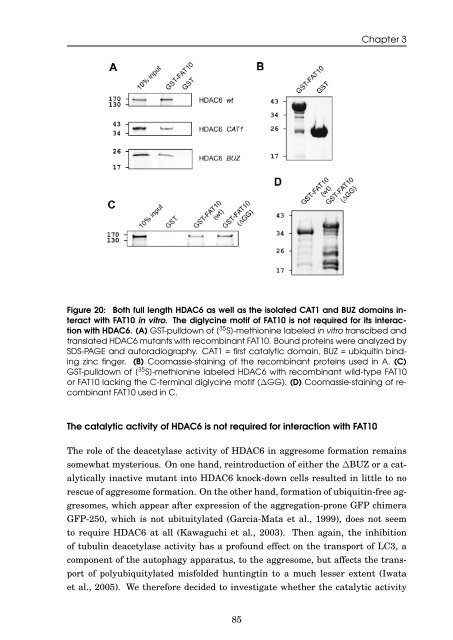
Figure 20: Both full length HDAC6 as well as <strong>the</strong> isolated CAT1 <strong>and</strong> BUZ doma<strong>in</strong>s <strong>in</strong>teract<br />
with <strong>FAT10</strong> <strong>in</strong> vitro. The diglyc<strong>in</strong>e motif <strong>of</strong> <strong>FAT10</strong> is not required for its <strong>in</strong>teraction<br />
with HDAC6. (A) GST-pulldown <strong>of</strong> [ 35 S]-methion<strong>in</strong>e labeled <strong>in</strong> vitro transcibed <strong>and</strong><br />
translated HDAC6 mutants with recomb<strong>in</strong>ant <strong>FAT10</strong>. Bound prote<strong>in</strong>s were analyzed by<br />
SDS-PAGE <strong>and</strong> autoradiography. CAT1 = first catalytic doma<strong>in</strong>, BUZ = <strong>ubiquit<strong>in</strong></strong> b<strong>in</strong>d<strong>in</strong>g<br />
z<strong>in</strong>c f<strong>in</strong>ger. (B) Coomassie-sta<strong>in</strong><strong>in</strong>g <strong>of</strong> <strong>the</strong> recomb<strong>in</strong>ant prote<strong>in</strong>s used <strong>in</strong> A. (C)<br />
GST-pulldown <strong>of</strong> [ 35 S]-methion<strong>in</strong>e labeled HDAC6 with recomb<strong>in</strong>ant wild-type <strong>FAT10</strong><br />
or <strong>FAT10</strong> lack<strong>in</strong>g <strong>the</strong> C-term<strong>in</strong>al diglyc<strong>in</strong>e motif (∆GG). (D) Coomassie-sta<strong>in</strong><strong>in</strong>g <strong>of</strong> recomb<strong>in</strong>ant<br />
<strong>FAT10</strong> used <strong>in</strong> C.<br />
The catalytic activity <strong>of</strong> HDAC6 is not required for <strong>in</strong>teraction with <strong>FAT10</strong><br />
The role <strong>of</strong> <strong>the</strong> deacetylase activity <strong>of</strong> HDAC6 <strong>in</strong> aggresome formation rema<strong>in</strong>s<br />
somewhat mysterious. On one h<strong>and</strong>, re<strong>in</strong>troduction <strong>of</strong> ei<strong>the</strong>r <strong>the</strong> ∆BUZ or a cat-<br />
alytically <strong>in</strong>active mutant <strong>in</strong>to HDAC6 knock-down cells resulted <strong>in</strong> little to no<br />
rescue <strong>of</strong> aggresome formation. On <strong>the</strong> o<strong>the</strong>r h<strong>and</strong>, formation <strong>of</strong> <strong>ubiquit<strong>in</strong></strong>-free ag-<br />
gresomes, which appear after expression <strong>of</strong> <strong>the</strong> aggregation-prone GFP chimera<br />
GFP-250, which is not ubituitylated (Garcia-Mata et al., 1999), does not seem<br />
to require HDAC6 at all (Kawaguchi et al., 2003). Then aga<strong>in</strong>, <strong>the</strong> <strong>in</strong>hibition<br />
<strong>of</strong> tubul<strong>in</strong> deacetylase activity has a pr<strong>of</strong>ound effect on <strong>the</strong> transport <strong>of</strong> LC3, a<br />
component <strong>of</strong> <strong>the</strong> autophagy apparatus, to <strong>the</strong> aggresome, but affects <strong>the</strong> trans-<br />
port <strong>of</strong> polyubiquitylated misfolded hunt<strong>in</strong>gt<strong>in</strong> to a much lesser extent (Iwata<br />
et al., 2005). We <strong>the</strong>refore decided to <strong>in</strong>vestigate whe<strong>the</strong>r <strong>the</strong> catalytic activity<br />
85














