

liiiMIIIfl~UDliiiMIII~U - Biblioteca de la Universidad Complutense ...
liiiMIIIfl~UDliiiMIII~U - Biblioteca de la Universidad Complutense ...
liiiMIIIfl~UDliiiMIII~U - Biblioteca de la Universidad Complutense ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Canítulo 1 Introducción<br />
en forma <strong>de</strong> complejos con <strong>la</strong> albúmina, son oxidados<br />
por el hígado y los tejidos extrahepáticos<br />
(Sug<strong>de</strong>n et al., 1989; Eeylot, 1996). En situaciones<br />
como ésta, en <strong>la</strong>s que <strong>la</strong> oxidación <strong>de</strong> ácidos grasos<br />
se encuentra estimu<strong>la</strong>da, parecen una serie <strong>de</strong> mecanismos<br />
adaptativos que conducen a <strong>la</strong> supresión<br />
<strong>de</strong> <strong>la</strong> utilización y oxidación <strong>de</strong> <strong>la</strong> glucosa (el ciclo<br />
glucosa/ácidos grasos), por lo que los requerimientos<br />
gluconeogénicos para el mantenimiento <strong>de</strong><br />
<strong>la</strong> glucemia disminuyen (Fig 1). Así, <strong>la</strong> oxidación<br />
<strong>de</strong> ácidos grasos inhibe <strong>la</strong> fosfori<strong>la</strong>ción <strong>de</strong> <strong>la</strong> glucosa,<br />
el flujo glicolítico a nivel <strong>de</strong> <strong>la</strong> 6-fosfofructo-<br />
1-quinasa, <strong>la</strong> enzima regu<strong>la</strong>dora <strong>de</strong> <strong>la</strong> primera fase<br />
<strong>de</strong> <strong>la</strong> glicolisis y <strong>la</strong> oxidación <strong>de</strong> piruvato por el<br />
complejo <strong>de</strong> <strong>la</strong> piruvato <strong>de</strong>shidrogenasa (Hue el al,<br />
1988). Este último efecto permite que se reciclen<br />
los esqueletos <strong>de</strong> carbono entre <strong>la</strong> glicolisis y <strong>la</strong><br />
gluconeogénesis (ciclo <strong>de</strong> Cori, Fig. 1). La oxidación<br />
<strong>de</strong> ácidos grasos estimu<strong>la</strong> a<strong>de</strong>más <strong>la</strong> gluconeogénesis<br />
hepática (Sug<strong>de</strong>n el aL, 1989). En<br />
suma, <strong>la</strong> oxidación <strong>de</strong> <strong>la</strong>s reservas lipídicas juega<br />
un papel regu<strong>la</strong>dor <strong>de</strong> gran importancia tanto en <strong>la</strong><br />
CC AG-Ab TO<br />
CC t Ins/Gluc (ca<strong>de</strong>na <strong>la</strong>rga)<br />
conservación como en <strong>la</strong> biosíntesis <strong>de</strong> <strong>la</strong> glucosa.<br />
En situaciones <strong>de</strong> ingesta correcta, una parte<br />
<strong>de</strong> los ácidos grasos hepáticos se <strong>de</strong>svía hacia <strong>la</strong><br />
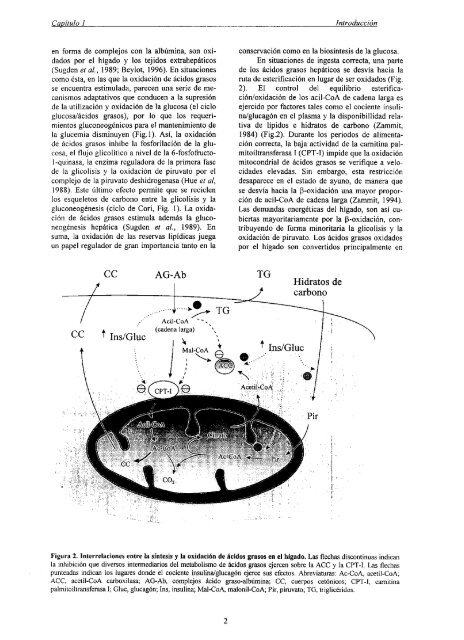
ruta <strong>de</strong> esterificación en lugar <strong>de</strong> ser oxidados (Fig.<br />
2). El control <strong>de</strong>l equilibrio esterificación/oxidación<br />
<strong>de</strong> los acil-CoA <strong>de</strong> ca<strong>de</strong>na <strong>la</strong>rga es<br />
ejercido por factores tales como el cociente insulina/glucagón<br />
en el p<strong>la</strong>sma y <strong>la</strong> disponibillidad re<strong>la</strong>tiva<br />
<strong>de</strong> lípidos e hidratos <strong>de</strong> carbono (Zammit,<br />
1984) (Fig.2). Durante los periodos <strong>de</strong> alimentación<br />
correcta, <strong>la</strong> baja actividad <strong>de</strong> <strong>la</strong> camitina palmitoiltransferasa<br />
1 (CPT-l) impi<strong>de</strong> que <strong>la</strong> oxidación<br />
mitocondrial <strong>de</strong> ácidos grasos se verifique a velocida<strong>de</strong>s<br />
elevadas. Sin embargo, esta restricción<br />
<strong>de</strong>saparece en el estado <strong>de</strong> ayuno, <strong>de</strong> manera que<br />
se <strong>de</strong>svía hacia <strong>la</strong> ~3-oxidaciónuna mayor proporción<br />
<strong>de</strong> acil-CoA <strong>de</strong> ca<strong>de</strong>na <strong>la</strong>rga (Zammit, 1994).<br />
Las <strong>de</strong>mandas energéticas <strong>de</strong>l hígado, son así cubiertas<br />
mayoritariamente por <strong>la</strong> j3-oxidación, contribuyendo<br />
<strong>de</strong> forma minoritaria <strong>la</strong> glicolisis y <strong>la</strong><br />
oxidación <strong>de</strong> piruvato. Los ácidos grasos oxidados<br />
por el hígado son convertidos principalmente en<br />
Acetil-.CoA<br />
Ins/GIuc<br />
Hidratos <strong>de</strong><br />
carbono<br />
Figura 2. Interre<strong>la</strong>ciones entre <strong>la</strong> síntesis y <strong>la</strong> oxidaci6n <strong>de</strong> ácidos grasos en el hígado. Las flechas discontinuas indican<br />
<strong>la</strong> inhibición que diversos intermediarios <strong>de</strong>l metabolismo <strong>de</strong> ácidos grasos ejercen sobre <strong>la</strong> ACC y <strong>la</strong> CPT-l. Las flechas<br />
punteadas indican los lugares don<strong>de</strong> el cociente insulina/glucagón ejerce sus efectos. Abreviaturas: Ac-CoA, acetil-CoA;<br />
ACC, acetil-CoA carboxi<strong>la</strong>sa; AO-Ab, complejos ácido graso-albúmina; CC, cuerpos cetónicos; CPT-l, camitina<br />
palmitoiltransferasa 1; Gluc, glucagón; ms, insulina; Mal-CoA, malonil-CoA; Pir, piruvato; TG, triglicéridos.<br />
2<br />
Pir<br />
1














