20 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 304 Ano<strong>the</strong>r possibility is that <strong>the</strong> Mayrellinae might have dispersed along <strong>the</strong> coasts <strong>of</strong> <strong>the</strong> Tethy Seaway during <strong>the</strong> early to middle Eocene. The Tethy Seaway was an important avenue <strong>of</strong> dispersal for <strong>the</strong> boreotropical flora across Eurasia, connecting eastern North America through western Europe to sou<strong>the</strong>astern Asia (Tiffney, 1985). Paleontological evidence reveals an Indomalaysian affinity <strong>of</strong> early Eocene to Oligocene floras <strong>of</strong> eastern Europe, Egypt, and England (London Clay) (Tiffney, 1985; Taylor, 1990, and references <strong>the</strong>rein). However, <strong>the</strong> existence <strong>of</strong> basal lineages <strong>of</strong> <strong>the</strong> Mayrellinae and Paramblynotus in <strong>the</strong> eastern Palearctic but not in <strong>the</strong> sou<strong>the</strong>astern Asia indicates that <strong>the</strong> Tethy Seaway was not <strong>the</strong> route <strong>of</strong> dispersal in this case. That <strong>the</strong> Bering land bridge is more likely to be <strong>the</strong> dispersal route for Mayrellinae between North America and eastern Palearctic/and Oriental is also supported by biogeographical reconstructions based on paleoclimatology and paleogeology. The Holarctic has been split and reconnected by various barriers at different locations since <strong>the</strong> early breakup <strong>of</strong> Pangaea in <strong>the</strong> Late Jurassic (e.g., Hallam, 1981; McKenna, 1983; Briggs, 1987). These events have pr<strong>of</strong>oundly affected <strong>the</strong> evolution <strong>of</strong> organisms <strong>of</strong> <strong>the</strong> Nor<strong>the</strong>rn Hemisphere (McKenna, 1983) as well as <strong>the</strong> present distribution patterns <strong>of</strong> <strong>the</strong>se organisms. These barriers are (1) <strong>the</strong> Turgai Sea, (2) <strong>the</strong> North <strong>American</strong> mid-continental seaway, (3) <strong>the</strong> nor<strong>the</strong>rn Atlantic Ocean, and (4) <strong>the</strong> Bering area (e.g., Noonan, 1986; Briggs, 1987; Engh<strong>of</strong>f, 1995). The Turgai Sea was formed in <strong>the</strong> Middle Jurassic (160 Ma) and separated Asia from Europe (Hallam, 1981; Briggs, 1987). Species <strong>of</strong> Kiefferiella and Paramblynotus are only known to be associated with angiosperm trees. Although <strong>the</strong> angiosperms could have diverged before <strong>the</strong> Cretaceous (145 Ma) (Doyle and Donoghue; 1986, Crane et al., 1995), and <strong>the</strong> earliest angiosperm fossils are from <strong>the</strong> Valanginian (140–130 Ma) (Crane et al., 1995), angiosperms were generally lacking in <strong>the</strong> early Cretaceous (Wing and Sue, 1992). Therefore, early connections between North <strong>American</strong> and Asian elements <strong>of</strong> <strong>the</strong> Mayrellinae through Europe should have occurred after <strong>the</strong> formation <strong>of</strong> <strong>the</strong> Turgai Sea. By <strong>the</strong> Oligocene (30 Ma), when <strong>the</strong> Turgai Sea retreated (Hallam, 1981, Briggs, 1987), no land connection existed between Europe and North America (Hallam, 1981; Scrivastava and Tapscott, 1986). Therefore, <strong>the</strong> early connections between eastern Asian and North <strong>American</strong> elements <strong>of</strong> <strong>the</strong> Mayrellinae are more likely to have been by way <strong>of</strong> <strong>the</strong> Bering land bridge than by way <strong>of</strong> Europe. In <strong>the</strong> nor<strong>the</strong>rn Pacific, <strong>the</strong> Bering land bridge connected Asia and North America continuously since at least <strong>the</strong> Late Cretaceous (Briggs, 1987), possibly earlier (Hallam, 1981; Hickey, 1981). However, <strong>the</strong> Bering area as a route <strong>of</strong> biotic exchanges was severely constrained by climate and daylight length because <strong>the</strong> area was <strong>the</strong>n situated closer to <strong>the</strong> North Pole than it is today, and it has since slowly and gradually moved southward to its current position (McKenna, 1983). None<strong>the</strong>less, <strong>the</strong> global climates were much warmer and equitable during <strong>the</strong> late Cretaceous and early to middle Tertiary than <strong>the</strong>y are today (Wolfe, 1978, 1980, 1987; Tiffney, 1985; Wing and Sues, 1992). Climates became particularly favorable during <strong>the</strong> early Tertiary warm intervals, when <strong>the</strong> Bering land bridge was primarily covered with broadleaved, deciduous forest, perhaps with a thin sou<strong>the</strong>rn fringe <strong>of</strong> evergreen, mega<strong>the</strong>rmal communities (Tiffney, 1985; Wolfe, 1985). This definitely made it possible for <strong>the</strong> early Mayrellinae to disperse between <strong>the</strong> two sides <strong>of</strong> <strong>the</strong> land bridge. Within <strong>the</strong> historical framework laid out above, plausible reconstructions <strong>of</strong> <strong>the</strong> early biogeography <strong>of</strong> <strong>the</strong> Mayrellinae based on <strong>the</strong> three competing alternative hypo<strong>the</strong>ses can be presented as outlined below. The first two reconstructions (figs. 16, 17) suggest that <strong>the</strong> ancestral species <strong>of</strong> Mayrellinae was distributed both in <strong>the</strong> Nearctic and in East Asia, and it was divided into disjunct populations when a geographical barrier separated <strong>the</strong>se two regions, giving rise to <strong>the</strong> genera Paramblynotus and Kiefferiella. The stem species <strong>of</strong> Paramblynotus expanded its distribution by dispersal into <strong>the</strong> Nearctic and became split by a subsequent vicariance event, leading to <strong>the</strong> origination <strong>of</strong> <strong>the</strong> P. virginianus clade and <strong>the</strong> clade
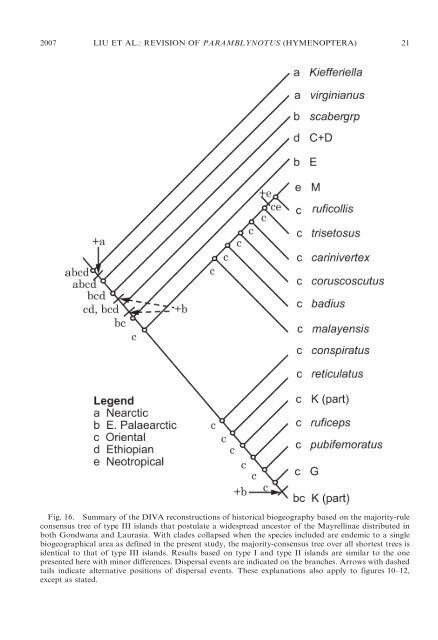
2007 LIU ET AL.: REVISION OF PARAMBLYNOTUS (HYMENOPTERA) 21 Fig. 16. Summary <strong>of</strong> <strong>the</strong> DIVA reconstructions <strong>of</strong> historical biogeography based on <strong>the</strong> majority-rule consensus tree <strong>of</strong> type III islands that postulate a widespread ancestor <strong>of</strong> <strong>the</strong> Mayrellinae distributed in both Gondwana and Laurasia. With clades collapsed when <strong>the</strong> species included are endemic to a single biogeographical area as defined in <strong>the</strong> present study, <strong>the</strong> majority-consensus tree over all shortest trees is identical to that <strong>of</strong> type III islands. Results based on type I and type II islands are similar to <strong>the</strong> one presented here with minor differences. Dispersal events are indicated on <strong>the</strong> branches. Arrows with dashed tails indicate alternative positions <strong>of</strong> dispersal events. These explanations also apply to figures 10–12, except as stated.