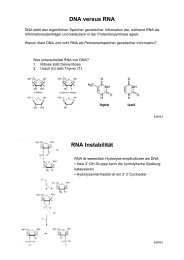
ZMBH J.Bericht 2000 - Zentrum für Molekulare Biologie der ...
ZMBH J.Bericht 2000 - Zentrum für Molekulare Biologie der ...
ZMBH J.Bericht 2000 - Zentrum für Molekulare Biologie der ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
TgMyoA: a candidate motor to power gliding<br />
motility<br />
C. Hettmann, A. Geiter , F, Delbac<br />
T. gondii myosin A (TgMyoA) is constitutively<br />
expressed and localizes beneath the plasma membrane<br />
of the parasite. Thus, TgMyoA is in an<br />
ideal position to transmit mechanical energy into<br />
forward motion propelling parasites into host cells.<br />
We mapped the membrane localization determinant<br />
within the short carboxy-terminal tail of TgMyoA,<br />
and site-directed mutagenesis revealed two essential<br />
arginine residues. The closest homologue in P. falciparum,<br />
PfMyoA is transcribed specifically in the<br />
invasive blood stage form and shows the same subcellular<br />
localization in merozoites. The nature of the<br />
receptor at the plasma membrane remains to be determined.<br />
TgMyoD and TgMyoE are similar to TgMyoA, but<br />
predominantly transcribed in the dormant bradyzoite<br />
stage. We have abrogated the expression of TgMyoD<br />
by double homologous recombination in the tachyzoites<br />
of the virulent RH strain without any apparent<br />
detectable phenotype. We need now to reproduce this<br />
knockout in a persistent strain capable to differentiate<br />
into bradyzoites, in or<strong>der</strong> to analyze the phenotypic<br />
consequences of the absence of TgMyoD in the<br />
appropriate stage.<br />
Biochemical characterization of TgMyoA<br />
A. Herm, in collaboration with M. Geeves (Kent<br />
UK), D. Manstein (MPI, Heidelberg) and E. Meyhöfer<br />
(Hannover)<br />
To power gliding motility, TgMyoA must be able to<br />
track along actin filaments upon ATP hydrolysis at<br />
a speed of 1-3 µm/s. We have determined the biochemical<br />
and physical properties of histidine tagged-<br />
132<br />
TgMyoA purified from culture recombinant parasites.<br />
Transient kinetic analysis of TgMyoA was<br />
obtained by stopped-flow and flash photolysis methods<br />
and revealed strong similarities to typical fast<br />
myosins, like rabbit skeletal myosin. The sliding<br />
velocity of fluorescently labeled actin filaments on<br />
TgMyoA attached to nitrocellulose-coated glass was<br />
5-6 µm/s. A step size of 5 to 6 nm was determined by<br />
single molecule assay monitoring filament displacement<br />
using optical tweezers. TgMyoA shows all the<br />
characteristics of a fast myosin of the class II but<br />
consi<strong>der</strong>ing the significant divergence in the converter<br />
domain and the quasi absence of neck, it is<br />
unclear at the moment how such a motor can generate<br />
a power stroke.<br />
TgMyoB/C: One gene, two tails, and two localizations.<br />
F. Delbac, C. Hettmann, A. Sänger<br />
TgMyoB and TgMyoC are alternative-spliced variants<br />
of the same gene, which produce two myosins<br />
exhibiting different tails. TgMyoB distributes evenly<br />
in the cytosol and is essentially soluble. In contrast,<br />
TgMyoC localizes at the posterior pole of the parasites<br />
describing a ring structure reminiscent of the<br />
termination of the inner membrane complex (IMC).<br />
TgMyoC protein fractionates with membranes and<br />
partitions in the detergent-insoluble fraction. Inhibitors<br />
of myosin ATPase and actin polymerization disrupt<br />
the or<strong>der</strong>ly turnover of mother cell organelles<br />
during daughter cell formation. The circular distribution<br />
of TgMyoC suggests a possible role of this<br />
myosin in the assembly and elongation of the IMCmicrotubule<br />
complex during parasite division. Moreover<br />
the overexpression of TgMyoB slows the rate of<br />
replication and leads to the formation of large resid-<br />
ual bodies, providing an additional indication of the<br />
involvement of TgMyoB/C in cell division.<br />
Figure 1. Immunolocalization of Myosin C (in red) and<br />
MIC6 (in green).<br />
II. Micronemal proteins of several apicomplexan<br />
parasites are sharing common<br />
structural and functional features<br />
The rapid and efficient nature of invasion by Apicomplexa<br />
relies on a sequence of events that are<br />
tightly controlled in time and space. Micronemes are<br />
involved in the trafficking and sequestration of binding<br />
ligands for host cell receptors. These organelles<br />
ensure the appropriate release of ligands in high concentration,<br />
at the tip of the parasite and upon response<br />
to external stimuli which senses contact with the<br />
host cells. To accommodate for the broad host range<br />
specificity of T. gondii, we postulated that adhesion<br />
should involve the recognition of ubiquitous surfaceexposed<br />
host molecules or, alternatively, the presence<br />
of various parasite attachment molecules able<br />
to recognize different host cell receptors. We identified<br />
and characterized a soluble protein MIC4, as<br />
well as a novel family of transmembrane microne-<br />
mal proteins in T. gondii. These proteins are released<br />
at the time of invasion and share domains of homology<br />
with proteins described in other members of the<br />
phylum Apicomplexa, supporting the hypothesis of a<br />
common molecular mechanism for host recognition<br />
attachment and invasion. They contain a combination<br />
of adhesive motifs including thrombospondin, integrin,<br />
apple and EGF-like domains. Invasion is successfully<br />
achieved when the newly formed parasitophorous<br />
vacuole is sealed and at this point, the<br />
tight interaction between host cell receptors and parasite<br />
ligands must be disrupted. The proteolytic processing<br />
of micronemal proteins that occurs on the<br />
parasite surface could fulfil such a function.<br />
TgMIC4 carries six apple domains and binds<br />
to host cells<br />
S. Brecht, U. Jäkle<br />
TgMIC4 carries six apple domains, a signal peptide<br />
at the amino terminus and no apparent membranespanning<br />
domain. Apple domains are found in plasma<br />
coagulation factors, kallikrein and factor XI and are<br />
involved in highly specific protein-protein interactions.<br />
MIC4 is expressed in all invasive stages of the<br />
parasites, secreted at the time of invasion and proteolytically<br />
processed twice onto the parasite surface,<br />
after secretion by the micronemes. MIC4 binds efficiently<br />
to host cells and the adhesive properties map<br />
within the carboxy-terminal apple domain of the protein.<br />
Preliminary experiments showed that galactose<br />
inhibits specifically binding to host cells and we are<br />
currently characterizing further the lectin properties<br />
of MIC4 and attempting to identify the host cell<br />
receptor(s) by expressing heterologously the apple<br />
domain in P. pastoris.<br />
133