Principles of cell signaling - UT Southwestern
Principles of cell signaling - UT Southwestern
Principles of cell signaling - UT Southwestern

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
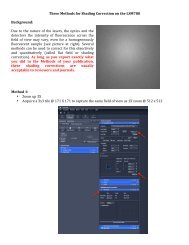
39057_ch14_<strong>cell</strong>bio.qxd 8/28/06 5:11 PM Page 603<br />
Adaptation in <strong>signaling</strong> is one <strong>of</strong> the best examples<br />
<strong>of</strong> biological homeostasis. The adaptability<br />
<strong>of</strong> <strong>cell</strong>ular <strong>signaling</strong> can be quite impressive.<br />
Cells commonly regulate their sensitivity to physiological<br />
stimuli over more than a 100-fold range,<br />
and the mammalian visual response can adapt to<br />
incoming light over a 107-fold range. This remarkable<br />
ability allows a photoreceptor <strong>cell</strong> to<br />
detect a single photon, and allows a person to<br />
read in both very dim light and intense sunlight.<br />
Adaptability is observed in bacteria, plants, fungi,<br />
and animals. Many <strong>of</strong> its properties are conserved<br />
throughout biology, although the most complex<br />
adaptive mechanisms are found in animals. The<br />
general mechanism for adaptation is the negative<br />
feedback loop, which biochemically samples<br />
the signal and controls the adaptive process.<br />
Adaptation varies with both the intensity and<br />
the duration <strong>of</strong> the incoming signal. Stronger or<br />
more persistent inputs tend to drive greater adaptive<br />
change and, <strong>of</strong>ten, adaptation that persists<br />
for a longer time. Cells can modulate adaptation<br />
in this way because adaptation is exerted by a<br />
succession <strong>of</strong> independent mechanisms, each with<br />
its own sensitivity and kinetic parameters.<br />
G protein pathways <strong>of</strong>fer ex<strong>cell</strong>ent examples<br />
<strong>of</strong> adaptation. FIGURE 14.11 shows that the earliest<br />
step in adaptation is receptor phosphorylation,<br />
which is catalyzed by G protein-coupled<br />
receptor kinases (GRKs) that selectively recognize<br />
the receptor’s ligand-activated conformation.<br />
Phosphorylation inhibits the receptor’s ability<br />
to stimulate G protein activation and also promotes<br />
binding <strong>of</strong> arrestin, a protein that further<br />
inhibits G protein activation. Moreover, arrestin<br />
binding primes receptors for endocytosis, which<br />
removes them from the <strong>cell</strong> surface. Endocytosis<br />
can also be the first step in receptor proteolysis.<br />
Along with these direct effects, many receptor<br />
genes display feedback inhibition <strong>of</strong> transcription,<br />
such that <strong>signaling</strong> by a receptor decreases<br />
its own expression.<br />
Stimulation thus causes multiple adaptive<br />
processes that range from immediate (phosphorylation,<br />
arrestin binding) through delayed (transcriptional<br />
regulation), and include both reversible<br />
and irreversible events. This array <strong>of</strong> adaptive<br />
events has been demonstrated for many G protein-coupled<br />
receptors, and many <strong>cell</strong>s may use<br />
all <strong>of</strong> them to control output from one receptor.<br />
The speed, extent, and reversibility <strong>of</strong> adaptation<br />
are selected by a <strong>cell</strong>’s developmental program.<br />
Cells can change their patterns <strong>of</strong> adaptation<br />
both qualitatively and quantitatively by altering<br />
the points in a pathway where feedback is initiated<br />
and exerted. In a linear pathway, changing<br />
Multiple adaptation processes occur after a stimulus<br />
Relative<br />
response<br />
Receptor phosphorylation<br />
Arrestin binding<br />
Receptor<br />
endocytosis<br />
Endosomal receptor<br />
degradation<br />
5 Receptor transcription<br />
inhibited<br />
0 1 10 100 1000<br />
Time (seconds)<br />
Agonist<br />
added<br />
Agonist<br />
binds<br />
1<br />
2<br />
Agonist<br />
GPCR<br />
G protein<br />
Receptor<br />
recycling<br />
Lysosome<br />
3<br />
G protein<br />
active<br />
4<br />
EFFECTORS<br />
CYTOPLASM<br />
4 Receptor<br />
degradation<br />
DNA<br />
GRK<br />
1 Receptor<br />
Arrestin<br />
phosphorylation<br />
2 Arrestin<br />
binding<br />
NUCLEUS<br />
Early<br />
endosome<br />
GPCR gene<br />
3 Receptor<br />
endocytosis<br />
5 Receptor<br />
transcription<br />
inhibited<br />
FIGURE 14.11 Multiple adaptation processes are invoked during a stimulus,<br />
and multiple nested mechanisms for adaptation are the rule. They are usually<br />
invoked sequentially according to the duration and intensity <strong>of</strong> the stimulus.<br />
For GPCRs, at least five desensitizing mechanisms are known, with others acting<br />
on the G protein and effectors.<br />
these points will alter the kinetics or extent <strong>of</strong><br />
adaptation (Figure 14.10). In branched pathways,<br />
changing these points can determine whether<br />
adaptation is unique to one input or is exerted<br />
for many similar inputs. If receptor activation triggers<br />
its desensitization directly, or if an event<br />
downstream on an unbranched pathway triggers<br />
desensitization, then only signals that initiate with<br />
that receptor will be altered. Receptor-selective<br />
adaptation is referred to as homologous adaptation<br />
(Figure 14.10).<br />
14.10 Cellular <strong>signaling</strong> is remarkably adaptive 603