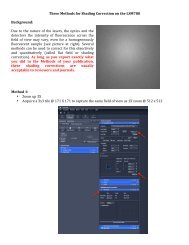
Principles of cell signaling - UT Southwestern
Principles of cell signaling - UT Southwestern
Principles of cell signaling - UT Southwestern

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
39057_ch14_<strong>cell</strong>bio.qxd 8/28/06 5:11 PM Page 618<br />
-strand repeats that form a cylindrical structure<br />
known as a propeller. There are five Gβ genes<br />
in mammals. Four encode strikingly similar proteins<br />
that naturally dimerize with the twelve Gγ<br />
subunits (Figure 14.24). The fifth, Gβ5, is less<br />
closely related to the others and interacts primarily<br />
with a Gγ-like domain in other proteins rather<br />
than with Gγ subunits themselves.<br />
Gγ subunits are smaller (~7 kDa) and far<br />
more diverse in sequence than are the Gβ’s. The<br />
last three amino acid residues <strong>of</strong> Gγ subunits<br />
are proteolyzed to leave a conserved C-terminal<br />
cysteine that is irreversibly S-prenylated<br />
and carboxymethylated, helping to anchor Gβγ<br />
to the membrane. Gβ and Gγ subunits can associate<br />
in most possible combinations. Because<br />
almost all <strong>cell</strong>s express multiple Gβ and Gγ subunits,<br />
it has been difficult to assign specific roles<br />
to individual Gβγ combinations. The best recognized<br />
interactions <strong>of</strong> Gβγ subunits occur at<br />
sites on Gβ, although distinct functions <strong>of</strong> Gγ<br />
have also been supported.<br />
14.21<br />
Heterotrimeric G proteins<br />
regulate a wide variety <strong>of</strong><br />
effectors<br />
Key concepts<br />
• G proteins convey signals by regulating the<br />
activities <strong>of</strong> multiple intra<strong>cell</strong>ular <strong>signaling</strong><br />
proteins known as effectors.<br />
• Effectors are structurally and functionally diverse.<br />
• A common G-protein binding domain has not been<br />
identified among effector proteins.<br />
• Effector proteins integrate signals from multiple G<br />
protein pathways.<br />
G protein-regulated effectors include enzymes<br />
that create or destroy intra<strong>cell</strong>ular second<br />
messengers (adenylyl cyclase, cyclic GMP phosphodiesterase,<br />
phospholipase C-β, phosphatidylinositol-3-kinase),<br />
protein kinases, ion channels<br />
(K+, Ca2+) and possibly membrane transport<br />
proteins (see Figure 14.25). Effectors may be<br />
integral membrane proteins or intrinsically soluble<br />
proteins that bind G proteins at the membrane<br />
surface. No conserved G protein-binding<br />
domain or sequence motif has been identified<br />
among effector proteins, and most effectors are<br />
related to proteins that have similar functions<br />
but that are not regulated by G proteins.<br />
Sensitivity to G protein regulation, thus, evolved<br />
independently in multiple families <strong>of</strong> regulatory<br />
proteins.<br />
Because they can respond to a variety <strong>of</strong><br />
Gα and Gβγ subunits, effector proteins can integrate<br />
signals from multiple G protein pathways.<br />
The different Gα or Gβγ subunits may<br />
have opposite or synergistic effects on a given<br />
effector. For example, some <strong>of</strong> the membranebound<br />
adenylyl cyclases in mammals are stimulated<br />
by Gα s<br />
and inhibited by Gα i<br />
(see Figure<br />
14.13). Many effectors are further regulated by<br />
other allosteric ligands (e.g., lipids, calmodulin)<br />
and by phosphorylation, contributing even more<br />
to integration <strong>of</strong> information.<br />
Effectors are usually represented as multiple<br />
is<strong>of</strong>orms, and each is<strong>of</strong>orm may be regulated<br />
differently, adding to the complexity <strong>of</strong> G<br />
protein networks. For example, some is<strong>of</strong>orms<br />
<strong>of</strong> adenylyl cyclase are stimulated by Gβγ,<br />
whereas others are inhibited. All phospholipase<br />
C-βs are stimulated both by Gα q<br />
family members<br />
and by Gβγ, but the potency and maximal<br />
effect <strong>of</strong> these two inputs vary dramatically<br />
among the four PLC-β is<strong>of</strong>orms.<br />
14.22<br />
Heterotrimeric G proteins<br />
are controlled by a<br />
regulatory GTPase cycle<br />
Key concepts<br />
• Heterotrimeric G proteins are activated when the<br />
Gα subunit binds GTP.<br />
• GTP hydrolysis to GDP inactivates the G protein.<br />
• GTP hydrolysis is slow, but is accelerated by<br />
proteins called GAPs.<br />
• Receptors promote activation by allowing GDP<br />
dissociation and GTP association; spontaneous<br />
exchange is very slow.<br />
• RGS proteins and phospholipase C-βs are GAPs for<br />
G proteins.<br />
The key event in heterotrimeric G protein <strong>signaling</strong><br />
is the binding <strong>of</strong> GTP to the Gα subunit.<br />
GTP binding activates the Gα subunit, which<br />
allows both it and the Gβγ subunit to bind and<br />
regulate effectors. The Gα subunit remains active<br />
as long as GTP is bound, but Gα also has<br />
GTPase activity and hydrolyzes bound GTP to<br />
GDP. Gα-GDP is inactive. G proteins thus traverse<br />
a GTPase cycle <strong>of</strong> GTP binding/activation and hydrolysis/deactivation,<br />
as depicted in FIGURE 14.26.<br />
Therefore, the control <strong>of</strong> G protein <strong>signaling</strong> is<br />
intrinsically kinetic. The relative signal strength,<br />
or amplitude, is proportional to the fraction <strong>of</strong><br />
G protein that is in the active, GTP-bound form.<br />
This fraction equals the balance <strong>of</strong> the rates <strong>of</strong><br />
GTP binding and GTP hydrolysis, the activating<br />
618 CHAPTER 14 <strong>Principles</strong> <strong>of</strong> <strong>cell</strong> <strong>signaling</strong>