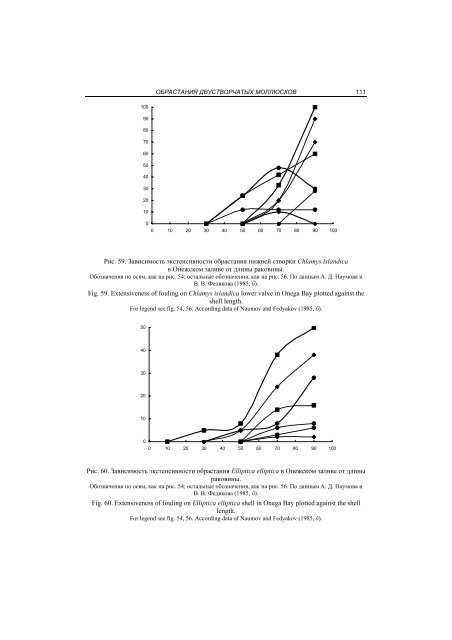
ОБРАСТАНИЯ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ 11110090807060504030201000 10 20 30 40 50 60 70 80 90 100Рис. 59. Зависимость экстенсивности обрастания нижней створки Chlamys islandicaв Онежском заливе от длины раковины.Обозначения по осям, как на рис. 54; остальные обозначения, как на рис. 56. По данным А. Д. Наумова иВ. В. Федякова (1985, б).Fig. 59. Extensiveness of fouling on Chlamys islandica lower valve in Onega Bay plotted against theshell length.For legend see fig. 54, 56. According data of Naumov and Fedyakov (1985, б).504030201000 10 20 30 40 50 60 70 80 90 100Рис. 60. Зависимость экстенсивности обрастания Elliptica elliptica в Онежском заливе от длиныраковины.Обозначения по осям, как на рис. 54; остальные обозначения, как на рис. 56. По данным А. Д. Наумова иВ. В. Федякова (1985, б).Fig. 60. Extensiveness of fouling on Elliptica elliptica shell in Onega Bay plotted against the shelllength.For legend see fig. 54, 56. According data of Naumov and Fedyakov (1985, б).
112Глава 6.ВЗАИМООТНОШЕНИЯ ДВУСТВОРОК И ДРУГИХ ОРГАНИЗМОВдам для этого требуется дорасти до 30% окончательной длины. На следующем этапек усоногим ракообразным присоединяются различные виды мшанок и, начиная с этогомомента, количество раковин, несущих на себе эти две группы обрастателей, остаетсяпримерно одинаковым. Именно они представляют собой наиболее массовыеформы организмов, поселяющихся на всех трех видах, причем крупные мидии и модиолюсыпрактически все несут на своих раковинах усоногих и мшанок, а Ellipticaelliptica – приблизительно в половине случаев.В развитии субдоминантных форм на раковинах этих трех видов моллюсков наблюдаютсянекоторые различия. Так, на Mytilus edulis по достижении ими половиныдефинитивного размера поселяются виды рода Heteranomia и Verruca stroemia, экстенсивностьобрастания которыми никогда не достигает 50%. Сходная картина наблюдаетсяв случае Elliptica elliptica с той разницей, что здесь роль субдоминантныхвидов играют асцидии (в основном это – Styelopsis grossularia) и Balanus. Иное делоModiolus modiolus, где второстепенные формы обрастателей, представленные видамирода Heteranomia и асцидиями (как правило, теми же, что и в предыдущем случае),появляются одновременно с доминирующими здесь Verruca stroemia. В дальнейшемасцидии входят в состав третьестепенных форм и только на самых крупных раковинахснова достигают той же экстенсивности, что и Heteranomia, заселяя их на 100%.Вообще из этих трех видов моллюсков Modiolus modiolus обладает наиболее богатойи разнообразной фауной обрастателей. На раковинах этого вида даже субдоминантныегруппы второго порядка встречаются в половине и более случаев. Это – асцидии,которые, как уже упоминалось, поселяются на всех крупных моллюсках,Chitinopoma fabricii, трубки которой встречены на трех четвертях особей максимальныхразмеров, и, наконец, виды родов Spirorbis и Balanus, экстенсивность седвазиикоторых достигает к концу жизни модиолуса 50%.Развитие обрастания Chlamys islandica довольно своеобразно, причем этот процесспротекает не одинаково на верхней и нижней створках (рис. 58–59). В первуюочередь отметим, что если на верхней створке первые седвазионты появляются подостижении моллюсками 10% окончательного размера, то на нижней – только 50%.Различия касаются и видового состава животных, поселяющихся на разных створкахгребешка: к верхней раньше всего прикрепляются различные мшанки, а к нижней –виды рода Spirorbis. Что же касается мшанок, то здесь они поселяются лишь по достижениираковиной 70% дефинитивной длины. Тем не менее крупные экземплярыChlamys islandica на три четверти обрастают мшанками. Это относится к обеимстворкам с той только разницей, что на верхней экстенсивность обрастания Bryozoaперестает возрастать, уже начиная с раковин, достигших половины окончательногоразмера, т. е. тогда, когда на нижней они еще не появились. Виды рода Spirorbis, доминирующиев начале на нижней стороне, на верхней оказываются в субдоминантномсостоянии. Экстенсивность обрастания ими здесь первое время возрастает, нопосле достижения моллюсками половины окончательных размеров эта группа начинаетуступать по обилию другим. Сходная динамика экстенсивности обрастания видамирода Spirorbis наблюдается и на нижней створке (см. рис. 59). Другие субдоминантныеформы обрастания верхней створки молодых гребешков – Chitinopoma fabriciiи асцидии (среди которых ведущее место занимает также Styelopsis grossularia) –имеют различную судьбу. Экстенсивность обрастания Chitinopoma fabricii возрастаетдо достижения моллюсками 70% максимального размера, а затем резко падает, чтонапоминает динамику, наблюдающуюся для видов рода Spirorbis. Что же касаетсяасцидий, то их экстенсивность довольно монотонно растет, хотя и не достигаетсколько-нибудь значительной величины, оставаясь в пределах 30%.По достижении раковинами половины окончательной длины картина несколькоменяется. Доминирующей группой все еще остаются мшанки. Субдоминантой перво-
- Page 2 and 3:
êéëëàâëäÄü ÄäÄÑÖåà
- Page 4 and 5:
êéëëàâëäÄü ÄäÄÑÖåà
- Page 6 and 7:
èêÖÑàëãéÇàÖДвуств
- Page 8 and 9:
ПРЕДИСЛОВИЕ 7мы «Ис
- Page 10 and 11:
É·‚‡ 1åÄíÖêàÄã à
- Page 12 and 13:
ДОННЫЕ ОРГАНИЗМЫ 11
- Page 14 and 15:
ДАННЫЕ ПО ГРУНТАМ 13
- Page 16 and 17:
ДАННЫЕ ПО ГРУНТАМ 15
- Page 18 and 19:
РАЗНООБРАЗИЕ И СТЕ
- Page 20 and 21:
ОЦЕНКА ОТРИЦАТЕЛЬН
- Page 22 and 23:
ВРЕМЕННЫЕ РЯДЫ 21ск
- Page 24 and 25:
РЕГРЕССИОННЫЙ АНАЛ
- Page 26 and 27:
РЕГРЕССИОННЫЙ АНАЛ
- Page 28 and 29:
РЕГРЕССИОННЫЙ АНАЛ
- Page 30 and 31:
АНАЛИЗ ЧИСЛА ЗУБОВ
- Page 32 and 33:
АНАЛИЗ ЧИСЛА ЗУБОВ
- Page 34 and 35:
ОСНОВНЫЕ ОБОЗНАЧЕН
- Page 36 and 37:
ОБЩАЯ ХАРАКТЕРИСТИ
- Page 38 and 39:
ГИДРОЛОГИЧЕСКИЕ ОС
- Page 40 and 41:
ФАУНИСТИЧЕСКАЯ ГРА
- Page 42 and 43:
ФАУНИСТИЧЕСКАЯ ГРА
- Page 44 and 45:
ФАУНИСТИЧЕСКАЯ ГРА
- Page 46 and 47:
ФАУНИСТИЧЕСКАЯ ГРА
- Page 48 and 49:
ФАУНИСТИЧЕСКАЯ ГРА
- Page 50 and 51:
ИСТОРИЯ БЕЛОГО МОР
- Page 52 and 53:
ИСТОРИЯ БЕЛОГО МОР
- Page 54 and 55:
ИСТОРИЯ БЕЛОГО МОР
- Page 56 and 57:
ИСТОРИЯ БЕЛОГО МОР
- Page 58 and 59:
ИСТОРИЯ БЕЛОГО МОР
- Page 60 and 61:
ИСТОРИЯ БЕЛОГО МОР
- Page 62 and 63: ИСТОРИЯ БЕЛОГО МОР
- Page 64 and 65: ИСТОРИЯ ИССЛЕДОВАН
- Page 66 and 67: ИСТОРИЯ ИССЛЕДОВАН
- Page 68 and 69: ИСТОРИЯ ИССЛЕДОВАН
- Page 70 and 71: СТЕПЕНЬ ИЗУЧЕННОСТ
- Page 72 and 73: СТЕПЕНЬ ИЗУЧЕННОСТ
- Page 74 and 75: СТЕПЕНЬ ИЗУЧЕННОСТ
- Page 76 and 77: СТЕПЕНЬ ИЗУЧЕННОСТ
- Page 78 and 79: СТЕПЕНЬ ИЗУЧЕННОСТ
- Page 80 and 81: СТЕПЕНЬ ИЗУЧЕННОСТ
- Page 82 and 83: É·‚‡ 5éëéÅÖççéëí
- Page 84 and 85: МНОГОЛЕТНЯЯ И СЕЗО
- Page 86 and 87: МНОГОЛЕТНЯЯ И СЕЗО
- Page 88 and 89: МНОГОЛЕТНЯЯ И СЕЗО
- Page 90 and 91: МНОГОЛЕТНЯЯ И СЕЗО
- Page 92 and 93: МОДЕЛЬ ДИНАМИКИ ОБ
- Page 94 and 95: МОДЕЛЬ ДИНАМИКИ ОБ
- Page 96: МОДЕЛЬ ДИНАМИКИ ОБ
- Page 99 and 100: 98Глава 5. ДИНАМИКА О
- Page 101 and 102: 100Глава 5. ДИНАМИКА
- Page 103 and 104: 102Глава 6. ВЗАИМООТН
- Page 105 and 106: 104Глава 6. ВЗАИМООТН
- Page 107 and 108: 106Глава 6.ВЗАИМООТН
- Page 109 and 110: 108Глава 6.ВЗАИМООТН
- Page 111: 110Глава 6.ВЗАИМООТН
- Page 115 and 116: 114Глава 6.ВЗАИМООТН
- Page 117 and 118: 116Глава 6.ВЗАИМООТН
- Page 119 and 120: 118Глава 6.ВЗАИМООТН
- Page 121 and 122: 120Глава 6.ВЗАИМООТН
- Page 123 and 124: É·‚‡ 7ÇÖêíàäÄãúç
- Page 125 and 126: 124Глава 7. ВЕРТИКАЛЬ
- Page 127 and 128: 126Глава 7. ВЕРТИКАЛЬ
- Page 129 and 130: 128Глава 7. ВЕРТИКАЛЬ
- Page 131 and 132: 130ГИДРОЛОГИЧЕСКИЕ
- Page 133 and 134: 132Глава 7. ВЕРТИКАЛЬ
- Page 135 and 136: 134Глава 7. ВЕРТИКАЛЬ
- Page 137 and 138: 136Глава 7. ВЕРТИКАЛЬ
- Page 139 and 140: 138Глава 7. ВЕРТИКАЛЬ
- Page 141 and 142: 140Глава 7. ВЕРТИКАЛЬ
- Page 143 and 144: 142Глава 7. ВЕРТИКАЛЬ
- Page 145 and 146: É·‚‡ 8éÅôÄü ïÄêÄ
- Page 147 and 148: 146Глава 8. ОБЩАЯ ХАР
- Page 149 and 150: 148Глава 8. ОБЩАЯ ХАР
- Page 151 and 152: 150Глава 8. ОБЩАЯ ХАР
- Page 153 and 154: 152Глава 8. ОБЩАЯ ХАР
- Page 155 and 156: 154Глава 8. ОБЩАЯ ХАР
- Page 157 and 158: 156Глава 8. ОБЩАЯ ХАР
- Page 159 and 160: 158Глава 9. ЗАСЕЛЕНИЕ
- Page 161 and 162: 160Глава 9. ЗАСЕЛЕНИЕ
- Page 163 and 164:
162Глава 9. ЗАСЕЛЕНИЕ
- Page 165 and 166:
164Глава 9. ЗАСЕЛЕНИЕ
- Page 167 and 168:
166Глава 9. ЗАСЕЛЕНИЕ
- Page 169 and 170:
168Глава 9. ЗАСЕЛЕНИЕ
- Page 171 and 172:
170Глава 9. ЗАСЕЛЕНИЕ
- Page 173 and 174:
É·‚‡ 10ëàëíÖåÄíà
- Page 175 and 176:
174Глава 10. СИСТЕМАТ
- Page 177 and 178:
176Глава 10. СИСТЕМАТ
- Page 179 and 180:
178Глава 10. СИСТЕМАТ
- Page 181 and 182:
180Глава 10. СИСТЕМАТ
- Page 183 and 184:
182Глава 10. СИСТЕМАТ
- Page 185 and 186:
184Глава 10. СИСТЕМАТ
- Page 187 and 188:
186Глава 10. СИСТЕМАТ
- Page 189 and 190:
188Глава 10. СИСТЕМАТ
- Page 191 and 192:
190Глава 10. СИСТЕМАТ
- Page 193 and 194:
192Глава 10. СИСТЕМАТ
- Page 195 and 196:
194Глава 10. СИСТЕМАТ
- Page 197 and 198:
196Глава 10. СИСТЕМАТ
- Page 199 and 200:
198Глава 10. СИСТЕМАТ
- Page 201 and 202:
200Глава 10. СИСТЕМАТ
- Page 203 and 204:
202Глава 10. СИСТЕМАТ
- Page 205 and 206:
204глава 10. СИСТЕМАТ
- Page 207 and 208:
206Глава 10. СИСТЕМАТ
- Page 209 and 210:
208Глава 10. СИСТЕМАТ
- Page 211 and 212:
210Глава 10. СИСТЕМАТ
- Page 213 and 214:
212Глава 10. СИСТЕМАТ
- Page 215 and 216:
214ГЛАВА 10. СИСТЕМАТ
- Page 217 and 218:
216ГЛАВА 10. СИСТЕМАТ
- Page 219 and 220:
218ГЛАВА 10. СИСТЕМАТ
- Page 221 and 222:
220Глава 10. СИСТЕМАТ
- Page 223 and 224:
222Глава 10. СИСТЕМАТ
- Page 225 and 226:
224Глава 10. СИСТЕМАТ
- Page 227 and 228:
226Глава 10. СИСТЕМАТ
- Page 229 and 230:
228Глава 10. СИСТЕМАТ
- Page 231 and 232:
230Глава 10. СИСТЕМАТ
- Page 233 and 234:
232Глава 10. СИСТЕМАТ
- Page 235 and 236:
234Глава 10. СИСТЕМАТ
- Page 237 and 238:
236Глава 10. СИСТЕМАТ
- Page 239 and 240:
238Глава 10. СИСТЕМАТ
- Page 241 and 242:
240Глава 10. СИСТЕМАТ
- Page 243 and 244:
242Глава 10. СИСТЕМАТ
- Page 245 and 246:
244Глава 10. СИСТЕМАТ
- Page 247 and 248:
246Глава 10. СИСТЕМАТ
- Page 249 and 250:
248Глава 10. СИСТЕМАТ
- Page 251 and 252:
250Глава 10. СИСТЕМАТ
- Page 253 and 254:
252Глава 10. СИСТЕМАТ
- Page 255 and 256:
254Глава 10. СИСТЕМАТ
- Page 257 and 258:
256Глава 10. СИСТЕМАТ
- Page 259 and 260:
258Глава 10. СИСТЕМАТ
- Page 261 and 262:
260Глава 10. СИСТЕМАТ
- Page 263 and 264:
262Глава 10. СИСТЕМАТ
- Page 265 and 266:
264Глава 10. СИСТЕМАТ
- Page 267 and 268:
266Глава 10. СИСТЕМАТ
- Page 269 and 270:
268Глава 10. СИСТЕМАТ
- Page 271 and 272:
270Глава 10. СИСТЕМАТ
- Page 273 and 274:
272Глава 10. СИСТЕМАТ
- Page 275 and 276:
274Глава 10. СИСТЕМАТ
- Page 277 and 278:
276Глава 10. СИСТЕМАТ
- Page 279 and 280:
278Глава 10. СИСТЕМАТ
- Page 281 and 282:
280Глава 10. СИСТЕМАТ
- Page 283 and 284:
282Глава 10. СИСТЕМАТ
- Page 285 and 286:
áÄäãûóÖçàÖДля выяс
- Page 287 and 288:
286ЗАКЛЮЧЕНИЕских ф
- Page 289 and 290:
288ЗАКЛЮЧЕНИЕкой и в
- Page 291 and 292:
290ЗАКЛЮЧЕНИЕчто эт
- Page 293 and 294:
SUMMARYA natural border should be f
- Page 295 and 296:
294SUMMARYis not cut down to partic
- Page 297 and 298:
296SUMMARYin the Gorlo Strait can p
- Page 299 and 300:
Приложение 1çÄïéÑä
- Page 301 and 302:
300ПРИЛОЖЕНИЕ 111. Muscul
- Page 303 and 304:
302ПРИЛОЖЕНИЕ 123. Nicani
- Page 305 and 306:
304ПРИЛОЖЕНИЕ 135. Arctic
- Page 307 and 308:
306ПРИЛОЖЕНИЕ 2Табли
- Page 309 and 310:
308ПРИЛОЖЕНИЕ 2Табли
- Page 311 and 312:
310ПРИЛОЖЕНИЕ 2Табли
- Page 313 and 314:
312ПРИЛОЖЕНИЕ 2Табли
- Page 315 and 316:
314ПРИЛОЖЕНИЕ 2Табли
- Page 317 and 318:
316ПРИЛОЖЕНИЕ 2Табли
- Page 319 and 320:
318ПРИЛОЖЕНИЕ 2Табли
- Page 321 and 322:
320ПРИЛОЖЕНИЕ 2Табли
- Page 323 and 324:
322ПРИЛОЖЕНИЕ 2Табли
- Page 325 and 326:
324ПРИЛОЖЕНИЕ 2Табли
- Page 327 and 328:
326ПРИЛОЖЕНИЕ 2Табли
- Page 329 and 330:
328ПРИЛОЖЕНИЕ 2Табли
- Page 331 and 332:
330ПРИЛОЖЕНИЕ 2Табли
- Page 333 and 334:
332ЛИТЕРАТУРАБеклем
- Page 335 and 336:
334ЛИТЕРАТУРАГурвич
- Page 337 and 338:
336ЛИТЕРАТУРАКолтун
- Page 339 and 340:
338ЛИТЕРАТУРАМаксим
- Page 341 and 342:
340ЛИТЕРАТУРАНаумов
- Page 343 and 344:
342ЛИТЕРАТУРАСемено
- Page 345 and 346:
344ЛИТЕРАТУРАAdams A. Des
- Page 347 and 348:
346ЛИТЕРАТУРАGalaktionov
- Page 349 and 350:
348ЛИТЕРАТУРАKnipowitsch
- Page 351 and 352:
350ЛИТЕРАТУРАReeve L. A.
- Page 353 and 354:
ÄíãÄëÅÖãéåéêëäàïÑ
- Page 367 and 368:
éÉãÄÇãÖçàÖПРЕДИСЛ
- Page 369 and 370:
CONTENTSPREFACE ...................
- Page 371:
Андрей Донатович Н