

Compendium of Potato Diseases - (PDF, 101 mb) - USAID
Compendium of Potato Diseases - (PDF, 101 mb) - USAID
Compendium of Potato Diseases - (PDF, 101 mb) - USAID

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
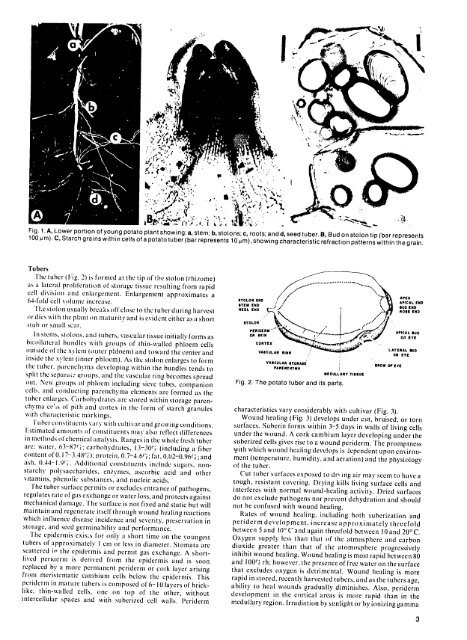
Fig. 1.A, Lower portion <strong>of</strong> young potato plant showing: a, stem; b, stolons; c, roots; and d, seed tuber. B, Bud on stolon tip (bar represents<br />
100 ,m). C, Starch grains within cells <strong>of</strong> a potato tuber (bar represents 10pm), showing characteristic refraction patterns within the grain.<br />
Tubers<br />
The tuber (Fig. 2) is formed at the tip <strong>of</strong> the stolon (rhizome)<br />
as a lateral proliferation <strong>of</strong> storage tissue resulting from rapid<br />
cell division and enlargement. Enlargement approximates a<br />
64-fold cell volme increase.<br />
'Re stolon usually breaks <strong>of</strong>fclose to the tuber during harvest<br />
or dies with the plant on maturity and is evident either as a short<br />
stub o r sm all scar .,E<br />
In stens, stolons, and tubers, vascular tissue initiallv forms as<br />
bikollateral bundles with groups <strong>of</strong> thin-walled phloem cells<br />
outside <strong>of</strong>'the xylem (outer phloem) and toward the center and<br />
inside the xyleml(inner phloem). As th' stolon enlarges to form<br />
the tuber. parenchyna developing within the bundles tends to<br />
split the separa it iroups. and the vascular ring becomes spread<br />
olt. New groups <strong>of</strong> phloem including sieve tubes, companion<br />
cells, and conducting parenchyma elements are formed as the<br />
tuber enlarges. Carbohydrates are stored within storage parenchyima<br />
ce',s <strong>of</strong> pith and cortex in the form <strong>of</strong> starch granules<br />
with characteristic markings.<br />
Tuber constitucnts vary with cultivar and growing conditions.<br />
E:stimated amounts <strong>of</strong> constituents may also reflect differences<br />
in methods <strong>of</strong>chemical analysis. Ranges in the whole fresh tuber<br />
are: water, 63-87i: carbohydrates, 13-30("i (including a fiber<br />
content <strong>of</strong>N. 17-3.48Ci ): protein, 0.7-4.61"; fat, 0.02-0.96(' ; and<br />
ash, 0.44-1.91". Additional constituents include sugars, notnstarchy<br />
polysaccharides, enzymes, ascorbic acid and other<br />
vitamins, phenolic substances, and nucleic acids,<br />
The tuber surface permits or excludes entrance <strong>of</strong> pathogens,<br />
regulates rate <strong>of</strong>gas exchange or water loss, and protects against<br />
mechanical damage. The surface is not fixed and static but will<br />
maintain and regenerate itself through wound healing reactions<br />
which influence disease incidence and severity, preservation in<br />
storage, and seed germinability and performance,<br />
Tle epidermis exists for only a short time on tile youngest<br />
tubers <strong>of</strong> approximately I cm or less in diameter. Stomata are<br />
scattered ir the epidermis and permit gas exchange. A shortlived<br />
penuerm is derived from tile epidermis and is soon<br />
replaced by a more permanent periderm or cork layer arising<br />
from meristenatic ca<strong>mb</strong>ium cells below the epidermis. This<br />
periderm in mature tubers is composed <strong>of</strong> 6-10 layers <strong>of</strong> bricklike,<br />
thin-wvalled cells, one on top <strong>of</strong> tie otiher, without<br />
intercellular spaces and with suberized cell walls. Periderm<br />
STEM ED J<br />
0<br />
\,\<br />
. ".<br />
" APEX<br />
BUDEND<br />
NEEL E, ROSE END<br />
STOLO<br />
,<br />
R ID, ' A C A L U D<br />
ORSK-N ' OR'eye<br />
CORTEX' LATERAL UD<br />
VASCULAR RING LAOR EYE<br />
VASCULAR ITORASE BO OFEY<br />
ARENCHYA !<br />
its Trts<br />
Fig. 2. The potato tuber and its parts.<br />
characteristics vary considerably with cultivar (Fig. 3).<br />
Wound healing (Fig. 3) develops under cut, bruised, or torn<br />
surfaces. Suberin forms within 3-5 days in walls <strong>of</strong> living cells<br />
under the wound. A cork ca<strong>mb</strong>ium layer developing under the<br />
suberized cells gives rise to a wound periderm. The promptness<br />
with which wound healing develops is Jependent upon environment<br />
(temperature, humidity, and aeration) and the physiology<br />
<strong>of</strong> the tuber.<br />
Cut tuber surfaces exposed to dr. ing air may seem to have a<br />
tough, resistant covering. Drying kills living surface cells and<br />
interferes with normal wound-healing activity. Dried surfaces<br />
do not exclude pathogens nor prevent dehydration and should<br />
not be confused with wound healing.<br />
Rates <strong>of</strong>' wound healing, including both suberization and<br />
periderm development, increase approximately threefold<br />
between 5 and 10' C and again threefold between 10 and 200 C.<br />
Oxygen supply less than that <strong>of</strong> the atmosphere and carbon<br />
dioxide greater than that <strong>of</strong> tile atomosphere progressively<br />
inhibit wound healing. Wound healing is most rapid between 80<br />
and 100%rh: however, the presence <strong>of</strong> free water on the surface<br />
that excludes oxygen is detrimental. Wound healing is more<br />
rapid in stored, recently harvested tubers, and as the tubers age,<br />
ability to heal wounds gradually diminishes. Also, periderm<br />
development in the cortical areas is more rapid than in the<br />
medullary region. Irradiation by sunlight or by ionizinggamma<br />
3














