

Principles of terrestrial ecosystem ecology.pdf
Principles of terrestrial ecosystem ecology.pdf
Principles of terrestrial ecosystem ecology.pdf

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
acids are metabolized more readily than cellulose<br />
and proteins, respectively. (2) Some chemical<br />
bonds are easier to break than others. Ester<br />
linkages that bind phosphate to organic skeletons<br />
and peptide bonds that link amino acids to<br />
form proteins, for example, are easier to break<br />
than the double bonds <strong>of</strong> aromatic rings. For<br />
these reasons, the nitrogen in proteins is much<br />
more available to microbes than the nitrogen<br />
contained in aromatic rings. (3) Compounds<br />
like lignin that have a highly irregular structure<br />
do not fit the active sites <strong>of</strong> most enzymes,<br />
so they are broken down much more slowly<br />
than are compounds like cellulose, which<br />
consist <strong>of</strong> chains <strong>of</strong> regularly repeating glucose<br />
units. (4) Some soluble compounds such as phenolics<br />
and alkaloids are toxic and kill or reduce<br />
the activity <strong>of</strong> microbes that absorb them. (5)<br />
Organic nitrogen and phosphorus are the major<br />
sources <strong>of</strong> nutrients for supporting microbial<br />
growth, so organic matter, such as straw,<br />
that contains low concentrations <strong>of</strong> these elements<br />
may not provide sufficient nutrients to<br />
allow microbes to use fully the carbon present<br />
in the litter.<br />
All <strong>of</strong> these chemical properties influence<br />
decomposition, but their relative importance is<br />
not well understood. Nonetheless, any <strong>of</strong> these<br />
properties can serve as a predictor <strong>of</strong> decomposition<br />
rate because the properties tend to be<br />
strongly correlated with one another. The ratio<br />
<strong>of</strong> carbon concentration to nitrogen concentration<br />
(C:N ratio), for example, has frequently<br />
been used as an index <strong>of</strong> litter quality, because<br />
litter with a low C:N ratio (high nitrogen<br />
concentration) generally decomposes quickly<br />
(Enríquez et al. 1993, Gholz et al. 2000).<br />
However, neither the nitrogen concentration<br />
<strong>of</strong> the litter nor the nitrogen availability in the<br />
soil directly influences the decomposition rate<br />
in most natural <strong>ecosystem</strong>s (Haynes 1986,<br />
Prescott 1995, Prescott et al. 1999, Hobbie and<br />
Vitousek 2000); this suggests that C:N ratio is<br />
not the chemical property that directly controls<br />
decomposition in these <strong>ecosystem</strong>s. This contrasts<br />
with agricultural residues such as straw,<br />
which have a low nitrogen concentration and a<br />
high concentration <strong>of</strong> moderately labile carbon<br />
sources like cellulose and hemicellulose. Nitro-<br />
Factors Controlling Decomposition 165<br />
gen concentration appears to limit directly the<br />
decomposition rate <strong>of</strong> organic matter primarily<br />
when labile carbon substrates are available to<br />
support microbial growth (Haynes 1986). This<br />
is more likely to occur in the rhizosphere than<br />
in fresh litter. Under other circumstances,<br />
carbon lability rather than nitrogen may be<br />
the primary control over decomposition rate<br />
(Hobbie 2000). Despite our uncertainty <strong>of</strong> the<br />
mechanistic role <strong>of</strong> C:N ratio in decomposition,<br />
many biogeochemical models use this<br />
ratio as a predictor <strong>of</strong> decomposition rate when<br />
different <strong>ecosystem</strong> types are compared (see<br />
Chapter 9).<br />
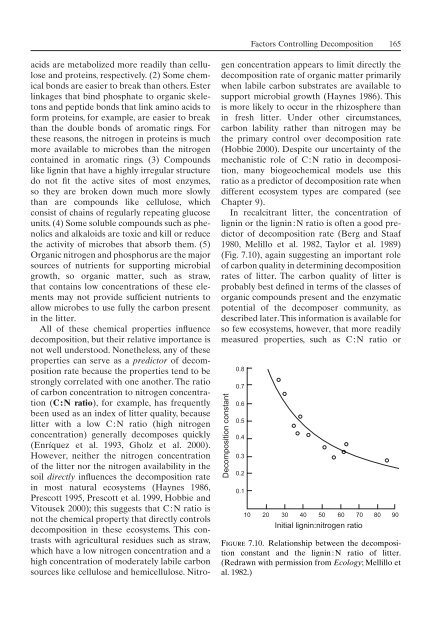
In recalcitrant litter, the concentration <strong>of</strong><br />
lignin or the lignin:N ratio is <strong>of</strong>ten a good predictor<br />
<strong>of</strong> decomposition rate (Berg and Staaf<br />
1980, Melillo et al. 1982, Taylor et al. 1989)<br />
(Fig. 7.10), again suggesting an important role<br />
<strong>of</strong> carbon quality in determining decomposition<br />
rates <strong>of</strong> litter. The carbon quality <strong>of</strong> litter is<br />
probably best defined in terms <strong>of</strong> the classes <strong>of</strong><br />
organic compounds present and the enzymatic<br />
potential <strong>of</strong> the decomposer community, as<br />
described later.This information is available for<br />
so few <strong>ecosystem</strong>s, however, that more readily<br />
measured properties, such as C:N ratio or<br />
Decomposition constant<br />
0.8<br />
0.7<br />
0.6<br />
0.5<br />
0.4<br />
0.3<br />
0.2<br />
0.1<br />
10 20 30 40 50 60 70 80 90<br />
Initial lignin:nitrogen ratio<br />
Figure 7.10. Relationship between the decomposition<br />
constant and the lignin : N ratio <strong>of</strong> litter.<br />
(Redrawn with permission from Ecology; Mellillo et<br />
al. 1982.)