

Principles of terrestrial ecosystem ecology.pdf
Principles of terrestrial ecosystem ecology.pdf
Principles of terrestrial ecosystem ecology.pdf

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
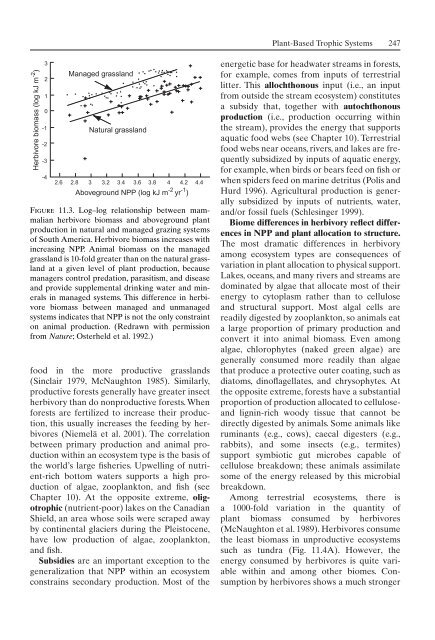
Herbivore biomass (log kJ m -2 )<br />
3<br />
2<br />
1<br />
0<br />
-1<br />
-2<br />
-3<br />
-4<br />
Managed grassland<br />
Natural grassland<br />
2.6 2.8 3 3.2 3.4 3.6 3.8 4 4.2 4.4<br />
Aboveground NPP (log kJ m -2 yr -1 )<br />
Figure 11.3. Log–log relationship between mammalian<br />
herbivore biomass and aboveground plant<br />
production in natural and managed grazing systems<br />
<strong>of</strong> South America. Herbivore biomass increases with<br />
increasing NPP. Animal biomass on the managed<br />
grassland is 10-fold greater than on the natural grassland<br />
at a given level <strong>of</strong> plant production, because<br />
managers control predation, parasitism, and disease<br />
and provide supplemental drinking water and minerals<br />
in managed systems. This difference in herbivore<br />
biomass between managed and unmanaged<br />
systems indicates that NPP is not the only constraint<br />
on animal production. (Redrawn with permission<br />
from Nature; Osterheld et al. 1992.)<br />
food in the more productive grasslands<br />
(Sinclair 1979, McNaughton 1985). Similarly,<br />
productive forests generally have greater insect<br />
herbivory than do nonproductive forests. When<br />
forests are fertilized to increase their production,<br />
this usually increases the feeding by herbivores<br />
(Niemelä et al. 2001). The correlation<br />
between primary production and animal production<br />
within an <strong>ecosystem</strong> type is the basis <strong>of</strong><br />
the world’s large fisheries. Upwelling <strong>of</strong> nutrient-rich<br />
bottom waters supports a high production<br />
<strong>of</strong> algae, zooplankton, and fish (see<br />
Chapter 10). At the opposite extreme, oligotrophic<br />
(nutrient-poor) lakes on the Canadian<br />
Shield, an area whose soils were scraped away<br />
by continental glaciers during the Pleistocene,<br />
have low production <strong>of</strong> algae, zooplankton,<br />
and fish.<br />
Subsidies are an important exception to the<br />
generalization that NPP within an <strong>ecosystem</strong><br />
constrains secondary production. Most <strong>of</strong> the<br />
Plant-Based Trophic Systems 247<br />
energetic base for headwater streams in forests,<br />
for example, comes from inputs <strong>of</strong> <strong>terrestrial</strong><br />
litter. This allochthonous input (i.e., an input<br />
from outside the stream <strong>ecosystem</strong>) constitutes<br />
a subsidy that, together with autochthonous<br />
production (i.e., production occurring within<br />
the stream), provides the energy that supports<br />
aquatic food webs (see Chapter 10). Terrestrial<br />
food webs near oceans, rivers, and lakes are frequently<br />
subsidized by inputs <strong>of</strong> aquatic energy,<br />
for example, when birds or bears feed on fish or<br />
when spiders feed on marine detritus (Polis and<br />
Hurd 1996). Agricultural production is generally<br />
subsidized by inputs <strong>of</strong> nutrients, water,<br />
and/or fossil fuels (Schlesinger 1999).<br />
Biome differences in herbivory reflect differences<br />
in NPP and plant allocation to structure.<br />
The most dramatic differences in herbivory<br />
among <strong>ecosystem</strong> types are consequences <strong>of</strong><br />
variation in plant allocation to physical support.<br />
Lakes, oceans, and many rivers and streams are<br />
dominated by algae that allocate most <strong>of</strong> their<br />
energy to cytoplasm rather than to cellulose<br />
and structural support. Most algal cells are<br />
readily digested by zooplankton, so animals eat<br />
a large proportion <strong>of</strong> primary production and<br />
convert it into animal biomass. Even among<br />
algae, chlorophytes (naked green algae) are<br />
generally consumed more readily than algae<br />
that produce a protective outer coating, such as<br />
diatoms, din<strong>of</strong>lagellates, and chrysophytes. At<br />
the opposite extreme, forests have a substantial<br />
proportion <strong>of</strong> production allocated to celluloseand<br />
lignin-rich woody tissue that cannot be<br />
directly digested by animals. Some animals like<br />
ruminants (e.g., cows), caecal digesters (e.g.,<br />
rabbits), and some insects (e.g., termites)<br />
support symbiotic gut microbes capable <strong>of</strong><br />
cellulose breakdown; these animals assimilate<br />
some <strong>of</strong> the energy released by this microbial<br />
breakdown.<br />
Among <strong>terrestrial</strong> <strong>ecosystem</strong>s, there is<br />
a 1000-fold variation in the quantity <strong>of</strong><br />
plant biomass consumed by herbivores<br />
(McNaughton et al. 1989). Herbivores consume<br />
the least biomass in unproductive <strong>ecosystem</strong>s<br />
such as tundra (Fig. 11.4A). However, the<br />
energy consumed by herbivores is quite variable<br />
within and among other biomes. Consumption<br />
by herbivores shows a much stronger