

Principles of terrestrial ecosystem ecology.pdf
Principles of terrestrial ecosystem ecology.pdf
Principles of terrestrial ecosystem ecology.pdf

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
188 8. Terrestrial Plant Nutrient Use<br />
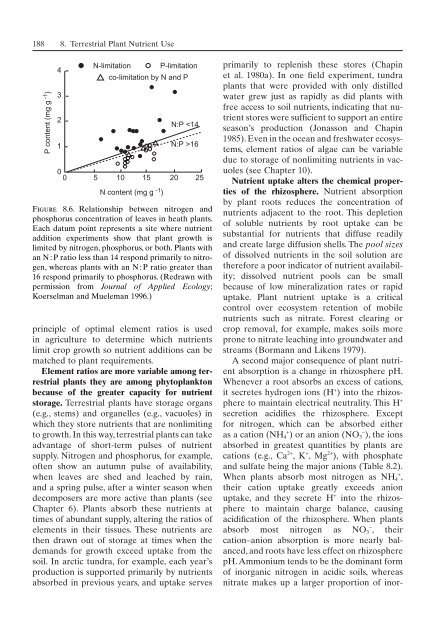
P content (mg g -1 )<br />
4<br />
3<br />
2<br />
1<br />
0 0<br />
N-limitation<br />
5<br />
10 15 20<br />
N content (mg g -1 )<br />
P-limitation<br />
co-limitation by N and P<br />
N:P 16<br />
Figure 8.6. Relationship between nitrogen and<br />
phosphorus concentration <strong>of</strong> leaves in heath plants.<br />
Each datum point represents a site where nutrient<br />
addition experiments show that plant growth is<br />
limited by nitrogen, phosphorus, or both. Plants with<br />
an N : P ratio less than 14 respond primarily to nitrogen,<br />
whereas plants with an N : P ratio greater than<br />
16 respond primarily to phosphorus. (Redrawn with<br />
permission from Journal <strong>of</strong> Applied Ecology;<br />
Koerselman and Mueleman 1996.)<br />
principle <strong>of</strong> optimal element ratios is used<br />
in agriculture to determine which nutrients<br />
limit crop growth so nutrient additions can be<br />
matched to plant requirements.<br />
Element ratios are more variable among <strong>terrestrial</strong><br />
plants they are among phytoplankton<br />
because <strong>of</strong> the greater capacity for nutrient<br />
storage. Terrestrial plants have storage organs<br />
(e.g., stems) and organelles (e.g., vacuoles) in<br />
which they store nutrients that are nonlimiting<br />
to growth. In this way, <strong>terrestrial</strong> plants can take<br />
advantage <strong>of</strong> short-term pulses <strong>of</strong> nutrient<br />
supply. Nitrogen and phosphorus, for example,<br />
<strong>of</strong>ten show an autumn pulse <strong>of</strong> availability,<br />
when leaves are shed and leached by rain,<br />
and a spring pulse, after a winter season when<br />
decomposers are more active than plants (see<br />
Chapter 6). Plants absorb these nutrients at<br />
times <strong>of</strong> abundant supply, altering the ratios <strong>of</strong><br />
elements in their tissues. These nutrients are<br />
then drawn out <strong>of</strong> storage at times when the<br />
demands for growth exceed uptake from the<br />
soil. In arctic tundra, for example, each year’s<br />
production is supported primarily by nutrients<br />
absorbed in previous years, and uptake serves<br />
25<br />
primarily to replenish these stores (Chapin<br />
et al. 1980a). In one field experiment, tundra<br />
plants that were provided with only distilled<br />
water grew just as rapidly as did plants with<br />
free access to soil nutrients, indicating that nutrient<br />
stores were sufficient to support an entire<br />
season’s production (Jonasson and Chapin<br />
1985). Even in the ocean and freshwater <strong>ecosystem</strong>s,<br />
element ratios <strong>of</strong> algae can be variable<br />
due to storage <strong>of</strong> nonlimiting nutrients in vacuoles<br />
(see Chapter 10).<br />
Nutrient uptake alters the chemical properties<br />
<strong>of</strong> the rhizosphere. Nutrient absorption<br />
by plant roots reduces the concentration <strong>of</strong><br />
nutrients adjacent to the root. This depletion<br />
<strong>of</strong> soluble nutrients by root uptake can be<br />
substantial for nutrients that diffuse readily<br />
and create large diffusion shells. The pool sizes<br />
<strong>of</strong> dissolved nutrients in the soil solution are<br />
therefore a poor indicator <strong>of</strong> nutrient availability;<br />
dissolved nutrient pools can be small<br />
because <strong>of</strong> low mineralization rates or rapid<br />
uptake. Plant nutrient uptake is a critical<br />
control over <strong>ecosystem</strong> retention <strong>of</strong> mobile<br />
nutrients such as nitrate. Forest clearing or<br />
crop removal, for example, makes soils more<br />
prone to nitrate leaching into groundwater and<br />
streams (Bormann and Likens 1979).<br />
A second major consequence <strong>of</strong> plant nutrient<br />
absorption is a change in rhizosphere pH.<br />
Whenever a root absorbs an excess <strong>of</strong> cations,<br />
it secretes hydrogen ions (H + ) into the rhizosphere<br />
to maintain electrical neutrality. This H +<br />
secretion acidifies the rhizosphere. Except<br />
for nitrogen, which can be absorbed either<br />
as a cation (NH4 + ) or an anion (NO3 - ), the ions<br />
absorbed in greatest quantities by plants are<br />
cations (e.g., Ca 2+ ,K + ,Mg 2+ ), with phosphate<br />
and sulfate being the major anions (Table 8.2).<br />
When plants absorb most nitrogen as NH4 + ,<br />
their cation uptake greatly exceeds anion<br />
uptake, and they secrete H + into the rhizosphere<br />
to maintain charge balance, causing<br />
acidification <strong>of</strong> the rhizosphere. When plants<br />
absorb most nitrogen as NO3 - , their<br />
cation–anion absorption is more nearly balanced,<br />
and roots have less effect on rhizosphere<br />
pH.Ammonium tends to be the dominant form<br />
<strong>of</strong> inorganic nitrogen in acidic soils, whereas<br />
nitrate makes up a larger proportion <strong>of</strong> inor-