

Inimese DNA koopiaarvu variatsioonid: nende tekkemehhanismid ja ...
Inimese DNA koopiaarvu variatsioonid: nende tekkemehhanismid ja ...
Inimese DNA koopiaarvu variatsioonid: nende tekkemehhanismid ja ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
mikrohomoloogiat lõhutud replikatsioonikahvli 3’ otsaga. Pakutakse, et MMBIR leiab aset<br />
siis, kui rakk on stressis. BIR-i toimumiseks on va<strong>ja</strong>lik Rad51 valkude olemasolu, mis omavad<br />
olulist funktsiooni <strong>DNA</strong> parandamisel, aktiveerides homoloogse rekombinatsiooni <strong>ja</strong> teisi<br />
mehhanisme, mis võtavad osa DSB reparatsioonist. Kui rakk on stressis, siis klassikaline BIR<br />
ei saa toimuda, kuna vastusena stressile on Rad51 valgud allaekspresseeritud ning<br />
homoloogsete interaktsioonide toimumise arv langeb, mille tulemusena BIR vahetakse<br />
MMBIR-i vastu. On näidanud, et näiteks hüpoksia viib Rad51 repressioonile <strong>ja</strong> seega kahaneb<br />
homoloogilise rekombinatsiooni sagedus (Hastings et al., 2009a).<br />
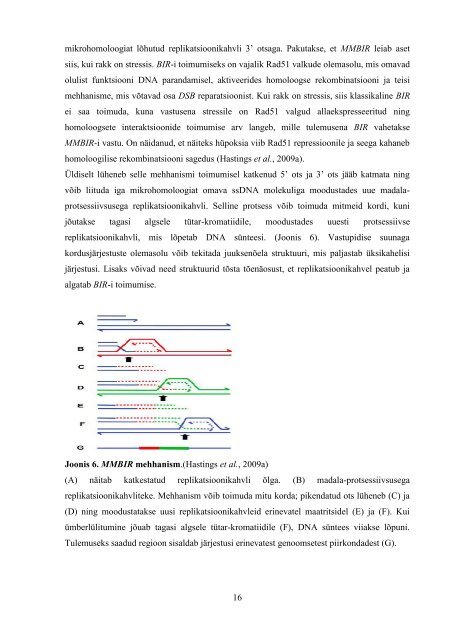
Üldiselt lüheneb selle mehhanismi toimumisel katkenud 5’ ots <strong>ja</strong> 3’ ots jääb katmata ning<br />
võib liituda iga mikrohomoloogiat omava ss<strong>DNA</strong> molekuliga moodustades uue madalaprotsessiivsusega<br />
replikatsioonikahvli. Selline protsess võib toimuda mitmeid kordi, kuni<br />
jõutakse tagasi algsele tütar-kromatiidile, moodustades uuesti protsessiivse<br />
replikatsioonikahvli, mis lõpetab <strong>DNA</strong> sünteesi. (Joonis 6). Vastupidise suunaga<br />
kordusjärjestuste olemasolu võib tekitada juuksenõela struktuuri, mis pal<strong>ja</strong>stab üksikahelisi<br />
järjestusi. Lisaks võivad need struktuurid tõsta tõenäosust, et replikatsioonikahvel peatub <strong>ja</strong><br />
algatab BIR-i toimumise.<br />
Joonis 6. MMBIR mehhanism.(Hastings et al., 2009a)<br />
(A) näitab katkestatud replikatsioonikahvli õlga. (B) madala-protsessiivsusega<br />
replikatsioonikahvliteke. Mehhanism võib toimuda mitu korda; pikendatud ots lüheneb (C) <strong>ja</strong><br />
(D) ning moodustatakse uusi replikatsioonikahvleid erinevatel maatritsidel (E) <strong>ja</strong> (F). Kui<br />
ümberlülitumine jõuab tagasi algsele tütar-kromatiidile (F), <strong>DNA</strong> süntees viiakse lõpuni.<br />
Tulemuseks saadud regioon sisaldab järjestusi erinevatest genoomsetest piirkondadest (G).<br />
16














