

Mechanisms of Olfaction in Insects - ResearchSpace@Auckland ...
Mechanisms of Olfaction in Insects - ResearchSpace@Auckland ...
Mechanisms of Olfaction in Insects - ResearchSpace@Auckland ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
General Introduction 20<br />
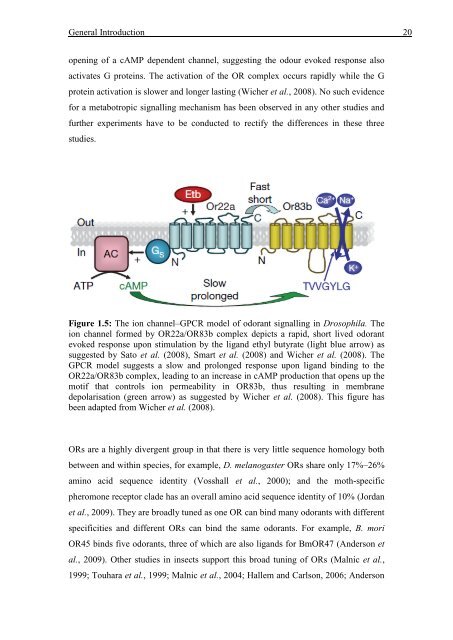
open<strong>in</strong>g <strong>of</strong> a cAMP dependent channel, suggest<strong>in</strong>g the odour evoked response also<br />
activates G prote<strong>in</strong>s. The activation <strong>of</strong> the OR complex occurs rapidly while the G<br />
prote<strong>in</strong> activation is slower and longer last<strong>in</strong>g (Wicher et al., 2008). No such evidence<br />
for a metabotropic signall<strong>in</strong>g mechanism has been observed <strong>in</strong> any other studies and<br />
further experiments have to be conducted to rectify the differences <strong>in</strong> these three<br />
studies.<br />
Figure 1.5: The ion channel–GPCR model <strong>of</strong> odorant signall<strong>in</strong>g <strong>in</strong> Drosophila. The<br />
ion channel formed by OR22a/OR83b complex depicts a rapid, short lived odorant<br />
evoked response upon stimulation by the ligand ethyl butyrate (light blue arrow) as<br />
suggested by Sato et al. (2008), Smart et al. (2008) and Wicher et al. (2008). The<br />
GPCR model suggests a slow and prolonged response upon ligand b<strong>in</strong>d<strong>in</strong>g to the<br />
OR22a/OR83b complex, lead<strong>in</strong>g to an <strong>in</strong>crease <strong>in</strong> cAMP production that opens up the<br />
motif that controls ion permeability <strong>in</strong> OR83b, thus result<strong>in</strong>g <strong>in</strong> membrane<br />
depolarisation (green arrow) as suggested by Wicher et al. (2008). This figure has<br />
been adapted from Wicher et al. (2008).<br />
ORs are a highly divergent group <strong>in</strong> that there is very little sequence homology both<br />
between and with<strong>in</strong> species, for example, D. melanogaster ORs share only 17%–26%<br />
am<strong>in</strong>o acid sequence identity (Vosshall et al., 2000); and the moth-specific<br />
pheromone receptor clade has an overall am<strong>in</strong>o acid sequence identity <strong>of</strong> 10% (Jordan<br />
et al., 2009). They are broadly tuned as one OR can b<strong>in</strong>d many odorants with different<br />
specificities and different ORs can b<strong>in</strong>d the same odorants. For example, B. mori<br />
OR45 b<strong>in</strong>ds five odorants, three <strong>of</strong> which are also ligands for BmOR47 (Anderson et<br />
al., 2009). Other studies <strong>in</strong> <strong>in</strong>sects support this broad tun<strong>in</strong>g <strong>of</strong> ORs (Malnic et al.,<br />
1999; Touhara et al., 1999; Malnic et al., 2004; Hallem and Carlson, 2006; Anderson











