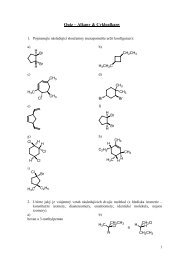
3. FOOD ChEMISTRy & bIOTEChNOLOGy 3.1. Lectures
3. FOOD ChEMISTRy & bIOTEChNOLOGy 3.1. Lectures
3. FOOD ChEMISTRy & bIOTEChNOLOGy 3.1. Lectures

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
Chem. Listy, 102, s265–s1311 (2008) Food Chemistry & Biotechnology<br />
P28 REACTION MEChANISM OF xyLOGLuCAN<br />
ENDOTRANSGLyCOSyLASE (xET) FROM<br />
PeTrOseliNuM CrisPuM ROOTS<br />
SOňA GARAJOVá, DAnA FLODROVá, VLADIMíR<br />
FARKAŠ and eVA STRATILOVá<br />
Institute of Chemistry, Slovak Academy of Sciences, Dúbravská<br />
cesta 9, 845 38 Bratislava, Slovakia<br />
chemsong@savba.sk<br />
Introduction<br />
Plant xyloglucan endotransglycosylases (XETs, EC<br />
2.4.1.207) catalyze the random cleavage of β-1,4-polyglucose<br />
backbone of the donor molecule (xyloglucan, XG) and<br />
the transfer of part of XG carrying the newly created reducing<br />
end to hydroxyl group at C-4 on the non-reducing end of<br />
another XG molecule or XG-derived oligosaccharide (acceptors).<br />
The process of transglycosylation can be described by<br />
Bi Bi reaction mechanism. Generally, there are two possible<br />
models of Bi Bi reaction mechanism: Pin-Pong and Sequential.<br />
In the Ping-Pong mechanism, the cleavage and transfer<br />
of part of polyglycan chain to the acceptor run in two steps.<br />
The first product (represented by the part of polyglycan chain<br />
carrying the original reducing end) is released from the stable<br />
enzyme-substrate complex before the acceptor substrate can<br />
bind. In contrast to the Ping-Pong mechanism, the sequential<br />
mechanism is observed like one step reaction, where a ternary<br />
complex of enzyme with the both substrates is formed.<br />
The products are released at the same time.<br />
The main XET form of parsley roots with the isoelectric<br />
point 4.6 has a broad pH optimum in the region of its stability<br />
(pH 4.5–9.0) with one maximum in acidic (pH 5.8) and<br />
the second one in alkalic (pH 8.8) region 1 . The kinetic analysis<br />
at these two pH optima was performed using radioactive<br />
alditols of XG octasaccharide (XLXGol + XXLGol) as<br />
an acceptor substrate as well as in dependence on the degree<br />
of polymerization (DP) of reducing xyloglucan oligosaccharides<br />
(XGOs). XGOs with DP 7, 8 and 9 were used.<br />
The mechanism of BiBi reaction was suggested from the<br />
nonlinear regression of kinetic data.<br />
Experimental<br />
E x t r a c t i o n o f X T H f r o m P a r s l e y<br />
R o o t s<br />
XET from parsley roots (Petroselinum crispum cv.<br />
Olomoucká dlouhá) was isolated and partially purified as<br />
described previously 1 .<br />
S u b s t r a t e s<br />
Tamarind seed xyloglucan used in this study was from<br />
Dainippon Pharmaceutical Co., Ltd, Osaka, Japan.<br />
XGOs with DP 7–9 were prepared by digestion of tamarind<br />
xyloglucan with Trichoderma cellulase 2 . They were<br />
purified on Biogel P2 column and further fractionated by<br />
preparative HPLC on TSK Gel Amide column (Tosoh) as<br />
described 3 . XG octasaccharide was converted to the corre-<br />
s633<br />
sponding 1-deoxy-1-aminoalditols (glycamines) by reductive<br />
amination 4 . Radioactive alditol of XG octasaccharide<br />
or reducing XGOs with DP 7, 8 and 9 were prepared by their<br />
reduction with na-borotritide [ 3 H]naBH 4 (ICn Radiochemicals,<br />
San Diego, CA) as described earlier 5 .<br />
K i n e t i c P a r a m e t e r s<br />
Standard XET assays were performed at 29 °C utilizing<br />
the radiometric method according to Fry et al. The initial<br />
rates were determined at pH 5.8 and 8.8 using five different<br />
concentrations of xyloglucan (0.5–5 g dm –3 at pH 5.8 and<br />
0.5–1.5 g dm –3 at pH 8.8, respectively) and of radioactive<br />
alditols of XG octasaccharide (0.6–<strong>3.</strong>1 µM). Further experiments<br />
were all performed at pH 5.8 using the same XG concentration<br />
range as described previously for this pH. Reducing<br />
XGOs with DP 7, 8 and 9 were used in the range of<br />
5–200 µM. The incorporation of each [1– 3 H]-labelled XGOs<br />
was measured by scintillation counting (Liquid Scintillation<br />
Analyzer Tri-Carb 2800TR, PerkinElmer, Illinois, USA) as<br />
described previously (Sulová et al., 1995). The K M values<br />
were calculated from nonlinear regression using Origin 6.0.<br />
Results<br />
XET as a member of glycoside hydrolase family GH16<br />
utilize a double displacement/retaining mechanism of transfer.<br />
This mechanism involves the formation of a covalent<br />
enzyme-substrate intermediate 6 , what is characteristic for<br />
1/v (nmol -1 .min)<br />
1/v (nmol -1 .min)<br />
75000<br />
50000<br />
25000<br />
150000<br />
100000<br />
50000<br />
0<br />
0 0.5 1 1.5 2 2.5<br />
1/XG (g -1 .L)<br />
0<br />
0 0.5 1 1.5 2 2.5<br />
1/XG (g -1 .L)<br />
Fig. 1. Plot of 1/v = f(1/XG) at different concentrations of mixture<br />
of XLXGol + XXLGol: 0.86 µM (▲), 1 µM (�), 1.24 µM (�),<br />
1.54 µM (◊), 2.1 µM (�) and <strong>3.</strong>1 µM (Δ). Michaelis parameters<br />
were determined at ph 5.8 (A) and 8.8 (b), respectively<br />
A<br />
B