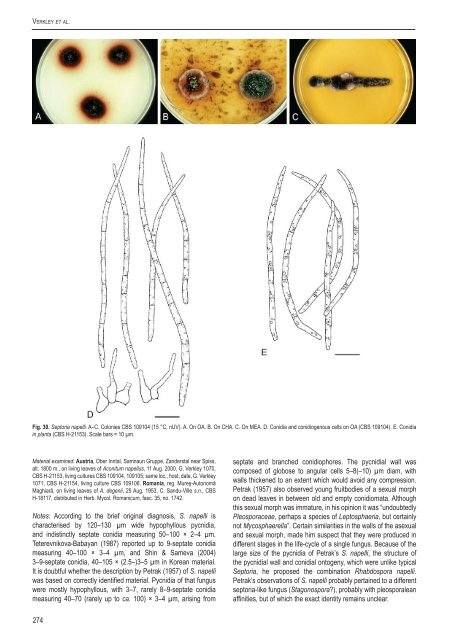
Verkley et al.Fig. 30. Sep<strong>to</strong>ria napelli. A–C. Colonies <strong>CBS</strong> 109104 (15 °C, nUV). A. On OA. B. On CHA. C. On MEA. D. Conidia and conidiogenous cells on OA (<strong>CBS</strong> 109104). E. Conidia<strong>in</strong> planta (<strong>CBS</strong> H-21153). Scale bars = 10 µm.Material exam<strong>in</strong>ed: Austria, Ober Inntal, Samnaun Gruppe, Zanderstal near Spiss,alt. 1800 m., on liv<strong>in</strong>g leaves of Aconitum napellus, 11 Aug. 2000, G. Verkley 1070,<strong>CBS</strong> H-21153, liv<strong>in</strong>g cultures <strong>CBS</strong> 109104, 109105; same loc., host, date, G. Verkley1071, <strong>CBS</strong> H-21154, liv<strong>in</strong>g culture <strong>CBS</strong> 109106. Romania, reg. Mureş-Au<strong>to</strong>nomăMaghiară, on liv<strong>in</strong>g leaves of A. degenii, 25 Aug. 1953, C. Sandu-Ville s.n., <strong>CBS</strong>H-18117, distributed <strong>in</strong> Herb. Mycol. Romanicum, fasc. 35, no. 1742.Notes: Accord<strong>in</strong>g <strong>to</strong> the brief orig<strong>in</strong>al diagnosis, S. napelli ischaracterised by 120–130 µm wide hypophyllous pycnidia,and <strong>in</strong>dist<strong>in</strong>ctly septate conidia measur<strong>in</strong>g 50–100 × 2–4 µm.Teterevnikova-Babayan (1987) reported up <strong>to</strong> 9-septate conidiameasur<strong>in</strong>g 40–100 × 3–4 µm, and Sh<strong>in</strong> & Sameva (2004)3–9-septate conidia, 40–105 × (2.5–)3–5 µm <strong>in</strong> Korean material.It is doubtful whether the description by Petrak (1957) of S. napelliwas based on correctly identified material. Pycnidia of that funguswere mostly hypophyllous, with 3–7, rarely 8–9-septate conidiameasur<strong>in</strong>g 40–70 (rarely up <strong>to</strong> ca. 100) × 3–4 µm, aris<strong>in</strong>g fromseptate and branched conidiophores. The pycnidial wall wascomposed of globose <strong>to</strong> angular cells 5–8(–10) µm diam, withwalls thickened <strong>to</strong> an extent which would avoid any compression.Petrak (1957) also observed young fruitbodies of a sexual morphon dead leaves <strong>in</strong> between old and empty conidiomata. Althoughthis sexual morph was immature, <strong>in</strong> his op<strong>in</strong>ion it was “undoubtedlyPleosporaceae, perhaps a <strong>species</strong> of Lep<strong>to</strong>sphaeria, but certa<strong>in</strong>lynot Mycosphaerella”. Certa<strong>in</strong> similarities <strong>in</strong> the walls of the asexualand sexual morph, made him suspect that they were produced <strong>in</strong>different stages <strong>in</strong> the life-cycle of a s<strong>in</strong>gle fungus. Because of thelarge size of the pycnidia of Petrak’s S. napelli, the structure ofthe pycnidial wall and conidial on<strong>to</strong>geny, which were unlike typicalSep<strong>to</strong>ria, he proposed the comb<strong>in</strong>ation Rhabdospora napelli.Petrak’s observations of S. napelli probably perta<strong>in</strong>ed <strong>to</strong> a differentsep<strong>to</strong>ria-like fungus (Stagonospora?), probably with pleosporaleanaff<strong>in</strong>ities, but of which the exact identity rema<strong>in</strong>s unclear.274
A <strong>new</strong> <strong>approach</strong> <strong>to</strong> <strong>species</strong> <strong>delimitation</strong> <strong>in</strong> Sep<strong>to</strong>riaThe fungus studied <strong>in</strong> the present study, which is a member ofthe Sep<strong>to</strong>ria clade, generally agrees with the orig<strong>in</strong>al descriptionof S. napelli. It is unknown whether S. napelli has a sexual morph.Two Mycosphaerella names have been published from Aconitum,M. an<strong>to</strong>novii on Aconitum excelsum <strong>in</strong> Siberia, and M. aconi<strong>to</strong>rum,on Aconitum sp. <strong>in</strong> Austria. Both names were <strong>in</strong>troduced byPetrak, who did not observe associated asexual morphs for theseMycosphaerella spp. A comparison with S. lycoc<strong>to</strong>ni, <strong>in</strong>clud<strong>in</strong>g themolecular results, is provide above <strong>in</strong> the notes on S. lycoc<strong>to</strong>ni.<strong>CBS</strong> 128664 isolated from Aconitum pseudolaeve var. erectum<strong>in</strong> Korea, is genetically dist<strong>in</strong>ct from both Sep<strong>to</strong>ria spp. on Aconitum<strong>in</strong> Europe. The <strong>new</strong> name S. pseudonapelli is proposed for thisfungus by Quaedvlieg et al. (2013, this volume).Sep<strong>to</strong>ria obesa Syd., <strong>in</strong> Syd. & P. Syd., Annls mycol. 12:163. 1914.= S. artemisiae Unamuno, Assoc. españ. Progr. Cienc. Congr. Salamanca: 46.1923 [nom. illeg., later homonym, non Passer<strong>in</strong>i, 1879].Descriptions <strong>in</strong> planta are provided by Punithal<strong>in</strong>gam (1967c) andPriest (2006). Sexual morph unknown.Hosts: Artemisia lavandulaefolia and Chrysanthemum spp.Material exam<strong>in</strong>ed: Germany, Weihenstephan, on Chrysanthemum <strong>in</strong>dicum, R.Schneider Sep. 1957, liv<strong>in</strong>g culture <strong>CBS</strong> 354.58 = BBA 8554 = IMI 091324. SouthKorea, Hongcheon, on Artemisia lavandulaefolia, H.D. Sh<strong>in</strong>, 28 June 2006, liv<strong>in</strong>gculture SMKC 21934 = KACC 42453 = <strong>CBS</strong> 128588; Bonghwa, on Chr. <strong>in</strong>dicum,H.D. Sh<strong>in</strong>, 18 Oct. 2007, liv<strong>in</strong>g culture SMKC 23048 = KACC 43193 = <strong>CBS</strong> 128623;Jeju, on Chr. morifolium, 5 July 2008, liv<strong>in</strong>g culture KACC 43858 = <strong>CBS</strong> 128759.Notes: Jørstad (1965) regarded S. obesa as a synonym of S.leucanthemi, as both have similar conidial morphologies andoccur on several Chrysanthemum spp. Punithal<strong>in</strong>gam (1967b, c),however, recognised S. obesa and S. leucanthemi as separate<strong>species</strong>, not<strong>in</strong>g that the conidia of S. obesa are consistently widerthan those of S. leucanthemi. Verkley & Star<strong>in</strong>k-Willemse (2004)found additional, molecular support for the treatment.as separate<strong>species</strong> <strong>in</strong> eight polymorphisms found on the ITS sequencesof stra<strong>in</strong>s represent<strong>in</strong>g these <strong>species</strong>. Further evidence is nowprovided here based on sequences of six other loci. The hostranges of the two <strong>species</strong> are also different: S. leucanthemiis capable of <strong>in</strong>fect<strong>in</strong>g various <strong>species</strong> of a wide range plantgenera, viz. Chrysanthemum, Tagetes, Achillea, Centaurea andHelianthus (Waddell & Weber 1963, Punithal<strong>in</strong>gam 1967b).Sep<strong>to</strong>ria obesa seems <strong>to</strong> ma<strong>in</strong>ly <strong>in</strong>fect Chrysanthemum spp.,but it does also <strong>in</strong>fect Artemisia lavendulaefolia, as could bedemonstrated <strong>in</strong> this study with <strong>CBS</strong> 128588, a stra<strong>in</strong> orig<strong>in</strong>allyidentified as S. artemisiae. The stra<strong>in</strong> is genetically very close <strong>to</strong>the other stra<strong>in</strong>s of S. obesa studied here and therefore regardedas conspecific. The conidia produced by <strong>CBS</strong> 128588 are <strong>in</strong> goodagreement with S. obesa as well, be<strong>in</strong>g much larger than <strong>in</strong> S.artemisiae (30–33 × 1.5 µm, accord<strong>in</strong>g <strong>to</strong> the orig<strong>in</strong>al diagnosisof S. artemisiae Passer<strong>in</strong>i). The later homonym S. artemisiaedescribed by Unamuno based on material on Artemisia vulgaris<strong>in</strong> Spa<strong>in</strong> with 4-septate conidia 35.5–52.5 × 2.5–3 µm, is placedhere <strong>in</strong> the synonymy of S. obesa.The conidia of the sunflower pathogen S. helianthi (50–85 ×2–3 μm) are similar <strong>to</strong> those of S. obesa (50–90 × 2.5–3.5 μm, cf.Priest 2006), but they can be dist<strong>in</strong>guished by the number of septaformed, viz., seldom more than 5 <strong>in</strong> S. helianthi and 5–11 septa <strong>in</strong>S. obesa. Verkley & Star<strong>in</strong>k already showed that ITS sequences ofthese <strong>species</strong> differ by more than 20 base positions, which is alsosupported by the results found <strong>in</strong> the present study for other genes(Fig. 2).Sep<strong>to</strong>ria paridis Pass., Atti Soc. crit<strong>to</strong>g. ital. 2: 41. 1879. Fig.31.Description <strong>in</strong> planta: Symp<strong>to</strong>ms leaf spots s<strong>in</strong>gle, scarce, circular <strong>to</strong>irregular, white <strong>to</strong> pale ochreous, surrounded by a vague orange <strong>to</strong>reddish brown zone, visible on both sides of the leaf, decay<strong>in</strong>g <strong>to</strong> shotholes.Conidiomata pycnidial, epiphyllous, one <strong>to</strong> a few <strong>in</strong> each leafspot, globose, black, immersed, 60–100 µm diam; ostiolum central,circular and 35–40 µm wide, surround<strong>in</strong>g cells concolorous <strong>to</strong> slightlydarker; conidiomatal wall up <strong>to</strong> 15 µm thick, composed throughout ofhyal<strong>in</strong>e, angular cells, 2.5–5 µm diam, the outermost cells brown withsomewhat thickened walls, the <strong>in</strong>ner cells hyal<strong>in</strong>e and th<strong>in</strong>-walled.Conidiogenous cells hyal<strong>in</strong>e, discrete, globose, doliiform, or broadlyampulliform, holoblastic, proliferat<strong>in</strong>g percurrently several times withdist<strong>in</strong>ct annellations thus form<strong>in</strong>g a relatively narrow neck, rarelyalso sympodially, 5–8(–11) × 2.5–5 µm. Conidia filiform, straight, orslightly curved, attenuated gradually <strong>to</strong> a narrowly po<strong>in</strong>ted apex anda narrowly truncate base, 0–3-septate (septa very th<strong>in</strong> and easilyoverlooked), not constricted around the septa, contents with severalm<strong>in</strong>ute oil-droplets and granular material <strong>in</strong> each cell <strong>in</strong> the liv<strong>in</strong>gstate, with m<strong>in</strong>ute oil-droplets and granular contents <strong>in</strong> the rehydratedstate, (18–)20–28.5(–34) × 1–1.5(–2) µm (liv<strong>in</strong>g; rehydrated, 1 µmwide). Sexual morph unknown.Description <strong>in</strong> vitro: Colonies on OA 8–11 mm diam <strong>in</strong> 10 d (30–35mm <strong>in</strong> 3 wk; more than 75 mm <strong>in</strong> 7 wk), with an even, glabrous,colourless marg<strong>in</strong>; immersed mycelium mostly homogeneouslypale coral <strong>to</strong> pale red, some pigment diffus<strong>in</strong>g beyond the colonymarg<strong>in</strong>, olivaceous <strong>to</strong> greenish hyphal radial strands also weaklyor more strongly develop<strong>in</strong>g <strong>in</strong> some sec<strong>to</strong>rs or entire colonies(especially after 7 wk, when most of the red pigment is no longervisible); <strong>in</strong> the centre olivaceous-black and slightly elevated due<strong>to</strong> superficial and immersed pycnidia, surrounded by an area withmore scattered pycnidia, releas<strong>in</strong>g pale whitish droplets of conidialslime; aerial mycelium very scanty, few m<strong>in</strong>ute white tufts; reverseolivaceous-black <strong>to</strong> greenish grey, surrounded by coral <strong>to</strong> siennaareas. Colonies on CMA 7–10 mm diam <strong>in</strong> 10 d (28–33 mm <strong>in</strong> 3wk; more than 75 mm <strong>in</strong> 7 wk), as on OA, but the colonies soonerpigmented, dark green, dark blueish green or olivaceous, and ared pigment tardily formed, but more persistent and still well visibleafter 7 wk. Sporulation as on OA. Colonies on MEA 6–11 mmdiam <strong>in</strong> 10 d (23–30 mm <strong>in</strong> 3 wk; 64–75 mm <strong>in</strong> 7 wk), the marg<strong>in</strong>even, glabrous, buff; colonies spread<strong>in</strong>g, but the centre elevated,irregularly pustulate, up <strong>to</strong> 2 mm high, the surface dark greyishbrown, later black, covered by short felty white aerial mycelium,or higher tufts; reverse of the colony brown-v<strong>in</strong>aceous or sepia,paler <strong>to</strong>wards the marg<strong>in</strong>. Pycnidia mostly superficial, <strong>in</strong> densegroups. Colonies on CHA 5–8 mm diam <strong>in</strong> 10 d (28–35 mm <strong>in</strong> 3 wk;45–55 mm <strong>in</strong> 7 wk), with an even <strong>to</strong> ruffled, glabrous, colourless <strong>to</strong>buff marg<strong>in</strong>; immersed mycelium <strong>in</strong> areas where first sporulationoccurs becom<strong>in</strong>g dark, greenish grey <strong>to</strong> dark slate blue, latermore throughout colony, covered by well-developed, tufty whitishgrey aerial mycelium that later shows a reddish haze; reverseolivaceous-black <strong>to</strong> sepia, but marg<strong>in</strong> paler; <strong>in</strong> the central part ofthe colony numerous pycnidia develop; <strong>in</strong> older colonies the centrebecomes up <strong>to</strong> 3 mm high.Conidiomata (OA) as <strong>in</strong> planta, immersed or develop<strong>in</strong>g onthe agar surface, s<strong>in</strong>gle or merged <strong>in</strong><strong>to</strong> complexes 100–220 µmwww.studies<strong>in</strong>mycology.org275