

Contents - Max-Planck-Institut für Physik komplexer Systeme
Contents - Max-Planck-Institut für Physik komplexer Systeme
Contents - Max-Planck-Institut für Physik komplexer Systeme

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
2.24 Pattern Formation in Active Fluids<br />
JUSTIN S. BOIS, FRANK JÜLICHER, STEPHAN W. GRILL<br />
Active stresses are exerted in cells and tissues by motor<br />
protein-driven contraction of the cytoskeleton. Concurrently,<br />
chemical species are reacting and diffusing<br />
within the material. Reaction-diffusion mechanisms<br />
have long been implicated in biological pattern formation<br />
of chemical species, but mechanical stresses are<br />
also crucial for developmental processes [1]. The active<br />
stress and distribution of chemical species associated<br />
with pattern formation are coupled in two ways. First,<br />
the active stress is under control of biochemical regulators<br />
affecting motor activity and actin polymerization,<br />
which may themselves diffuse and undergo chemical<br />
reactions. Secondly, the flow of contractile material<br />
transports chemical species by advection. In this<br />
work [2], we developed a new framework for biological<br />
pattern formation considering these two forms of<br />
coupling.<br />
In this first simplified treatment, we treat a contracting<br />
cytoskeleton or tissue as a thin film of an active viscous<br />
fluid and consider only movements in the x-direction.<br />
The equation of motion is then [3, 4]<br />
∂x ζ∆µ(c) = −η∂ 2 x v + γ v,<br />
where v is the fluid flow velocity, η is the viscosity, γ<br />
is a friction coefficient that takes into account boundary<br />
stresses, and ζ∆µ is the active stress, dependent on<br />
the concentrations of chemical species, c ≡ (c1,...,cn).<br />
The equation of motion reveals a physical length scale<br />
of the system, ℓ ≡ η/γ, which is the decay length of<br />
fluid velocity in response to a local active stress gradient<br />
[4]. In addition, mass conservation gives<br />
∂t ci = Di ∂ 2 x ci + Ri(c) − ∂x(ci v) ∀i ∈ [1,n],<br />
where Di is the diffusivity of species i and Ri(c) is the<br />
rate of production of species i by chemical reaction. The<br />
last term accounts for advective transport of species i<br />
that occurs by virtue of bulk fluid motion. The relevant<br />
dimensionless parameters are the ratio of diffusivities,<br />
Di/D1, and the Péclet number Pe ≡ (ζ∆µ)0 /γD1,<br />
where (ζ∆µ)0 is the maximal possible active stress.<br />
The Péclet number is the ratio of diffusive to advective<br />
times scales and is therefore a measure of the relative<br />
importance of advection in the system dynamics. Additionally,<br />
the functional forms of Ri(c) and ζ∆µ(c), as<br />
well as the dimensionless length of the system L/ℓ (the<br />
ratio of the total system extent to the hydrodynamic<br />
length), are important for the dynamics.<br />
Case study 1: active stress-driven instabilities. As<br />
a first case study, we consider a single biochemical<br />
species with concentration c that regulates active stress<br />
(a) (b)<br />
1<br />
0<br />
¢2¢1<br />
2£D¤(k)<br />
increasing<br />
Pe<br />
k¡<br />
0 1 2 3<br />
c 0<br />
10 2<br />
10 1<br />
10 0<br />
10 -1<br />
10 0 10 -2<br />
no patterns<br />
pattern-forming<br />
region<br />
10 1<br />
Pe/(1 +(¥¦¡/L) 2 )<br />
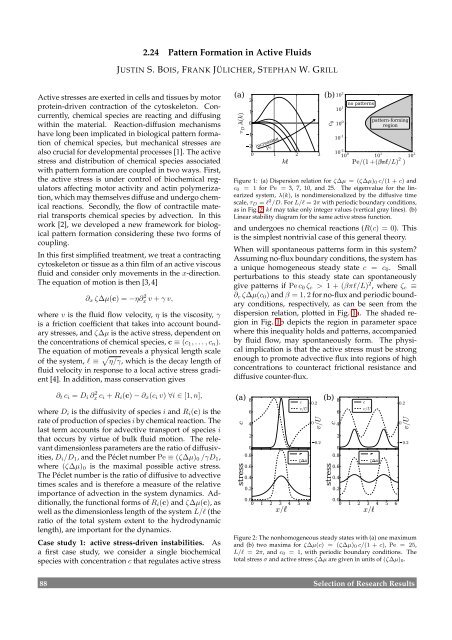
Figure 1: (a) Dispersion relation for ζ∆µ = (ζ∆µ)0 c/(1 + c) and<br />
c0 = 1 for Pe = 3, 7, 10, and 25. The eigenvalue for the linearized<br />
system, λ(k), is nondimensionalized by the diffusive time<br />
scale, τD = ℓ 2 /D. For L/ℓ = 2π with periodic boundary conditions,<br />
as in Fig. 2, kℓ may take only integer values (vertical gray lines). (b)<br />
Linear stability diagram for the same active stress function.<br />
and undergoes no chemical reactions (R(c) = 0). This<br />
is the simplest nontrivial case of this general theory.<br />
When will spontaneous patterns form in this system?<br />
Assuming no-flux boundary conditions, the system has<br />
a unique homogeneous steady state c = c0. Small<br />
perturbations to this steady state can spontaneously<br />
give patterns if Pe c0 ζc > 1 + (βπℓ/L) 2 , where ζc ≡<br />
∂c ζ∆µ(c0) and β = 1,2 for no-flux and periodic boundary<br />
conditions, respectively, as can be seen from the<br />
dispersion relation, plotted in Fig. 1a. The shaded region<br />
in Fig. 1b depicts the region in parameter space<br />
where this inequality holds and patterns, accompanied<br />
by fluid flow, may spontaneously form. The physical<br />
implication is that the active stress must be strong<br />
enough to promote advective flux into regions of high<br />
concentrations to counteract frictional resistance and<br />
diffusive counter-flux.<br />
(a)<br />
c<br />
stress<br />
8<br />
6<br />
4<br />
2<br />
0<br />
0.8<br />
0.6<br />
0.4<br />
0.2<br />
0 1 2 3 4 5<br />
x/§<br />
0.0<br />
6<br />
c<br />
v/U<br />
0.2<br />
v/U<br />
0<br />
(b)<br />
c<br />
2<br />
0.2 ¨©0 0.8<br />
0.6<br />
stress<br />
8<br />
6<br />
4<br />
0.4<br />
0.2<br />
c<br />
v/U<br />
0.2<br />
v/U<br />
0.2<br />
10 2<br />
¨©<br />
x/§<br />
0.0<br />
0 1 2 3 4 5 6<br />
Figure 2: The nonhomogeneous steady states with (a) one maximum<br />
and (b) two maxima for ζ∆µ(c) = (ζ∆µ)0 c/(1 + c), Pe = 25,<br />
L/ℓ = 2π, and c0 = 1, with periodic boundary conditions. The<br />
total stress σ and active stress ζ∆µ are given in units of (ζ∆µ)0.<br />
88 Selection of Research Results<br />
0













