

Growth, Differentiation and Sexuality
Growth, Differentiation and Sexuality
Growth, Differentiation and Sexuality

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
128 N.L. Glass <strong>and</strong> A. Fleissner<br />
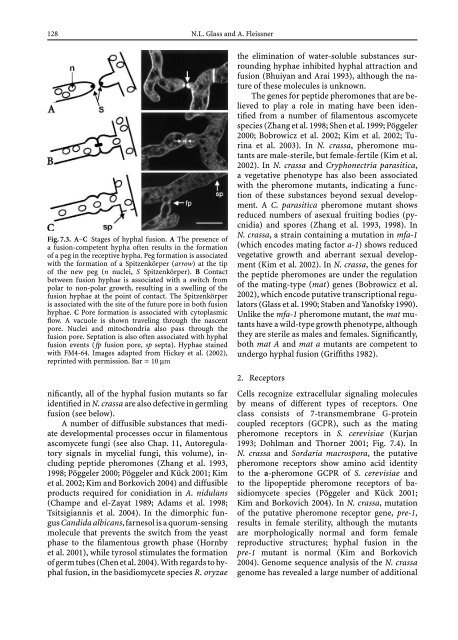
Fig. 7.3. A–C Stages of hyphal fusion. A The presence of<br />
a fusion-competent hypha often results in the formation<br />
of a peg in the receptive hypha. Peg formation is associated<br />
with the formation of a Spitzenkörper (arrow) atthetip<br />
of the new peg (n nuclei, S Spitzenkörper). B Contact<br />
between fusion hyphae is associated with a switch from<br />
polar to non-polar growth, resulting in a swelling of the<br />
fusion hyphae at the point of contact. The Spitzenkörper<br />
isassociatedwiththesiteofthefutureporeinbothfusion<br />
hyphae. C Pore formation is associated with cytoplasmic<br />
flow. A vacuole is shown traveling through the nascent<br />
pore. Nuclei <strong>and</strong> mitochondria also pass through the<br />
fusion pore. Septation is also often associated with hyphal<br />
fusion events (fp fusion pore, sp septa). Hyphae stained<br />
with FM4-64. Images adapted from Hickey et al. (2002),<br />
reprinted with permission. Bar = 10 μm<br />
nificantly, all of the hyphal fusion mutants so far<br />
identified in N. crassa are also defective in germling<br />
fusion (see below).<br />
A number of diffusible substances that mediate<br />
developmental processes occur in filamentous<br />
ascomycete fungi (see also Chap. 11, Autoregulatory<br />
signals in mycelial fungi, this volume), including<br />
peptide pheromones (Zhang et al. 1993,<br />
1998; Pöggeler 2000; Pöggeler <strong>and</strong> Kück 2001; Kim<br />
et al. 2002; Kim <strong>and</strong> Borkovich 2004) <strong>and</strong> diffusible<br />
products required for conidiation in A. nidulans<br />
(Champe <strong>and</strong> el-Zayat 1989; Adams et al. 1998;<br />
Tsitsigiannis et al. 2004). In the dimorphic fungusC<strong>and</strong>ida<br />
albicans, farnesol is a quorum-sensing<br />
molecule that prevents the switch from the yeast<br />
phase to the filamentous growth phase (Hornby<br />
et al. 2001), while tyrosol stimulates the formation<br />
of germ tubes (Chen et al. 2004). With regards to hyphal<br />
fusion, in the basidiomycete species R. oryzae<br />
the elimination of water-soluble substances surrounding<br />
hyphae inhibited hyphal attraction <strong>and</strong><br />
fusion (Bhuiyan <strong>and</strong> Arai 1993), although the nature<br />
of these molecules is unknown.<br />
The genes for peptide pheromones that are believedtoplayaroleinmatinghavebeenidentified<br />
from a number of filamentous ascomycete<br />
species (Zhang et al. 1998; Shen et al. 1999; Pöggeler<br />
2000; Bobrowicz et al. 2002; Kim et al. 2002; Turina<br />
et al. 2003). In N. crassa, pheromonemutants<br />
are male-sterile, but female-fertile (Kim et al.<br />
2002). In N. crassa <strong>and</strong> Cryphonectria parasitica,<br />
a vegetative phenotype has also been associated<br />
with the pheromone mutants, indicating a function<br />
of these substances beyond sexual development.<br />
A C. parasitica pheromone mutant shows<br />
reduced numbers of asexual fruiting bodies (pycnidia)<br />
<strong>and</strong> spores (Zhang et al. 1993, 1998). In<br />
N. crassa, a strain containing a mutation in mfa-1<br />
(which encodes mating factor a-1) showsreduced<br />
vegetative growth <strong>and</strong> aberrant sexual development<br />
(Kim et al. 2002). In N. crassa, thegenesfor<br />
the peptide pheromones are under the regulation<br />
of the mating-type (mat) genes (Bobrowicz et al.<br />
2002), which encode putative transcriptional regulators<br />
(Glass et al. 1990; Staben <strong>and</strong> Yanofsky 1990).<br />
Unlike the mfa-1 pheromone mutant, the mat mutants<br />
have a wild-type growth phenotype, although<br />
they are sterile as males <strong>and</strong> females. Significantly,<br />
both mat A <strong>and</strong> mat a mutants are competent to<br />
undergo hyphal fusion (Griffiths 1982).<br />
2. Receptors<br />
Cells recognize extracellular signaling molecules<br />
by means of different types of receptors. One<br />
class consists of 7-transmembrane G-protein<br />
coupled receptors (GCPR), such as the mating<br />
pheromone receptors in S. cerevisiae (Kurjan<br />
1993; Dohlman <strong>and</strong> Thorner 2001; Fig. 7.4). In<br />
N. crassa <strong>and</strong> Sordaria macrospora, the putative<br />
pheromone receptors show amino acid identity<br />
to the a-pheromone GCPR of S. cerevisiae <strong>and</strong><br />
to the lipopeptide pheromone receptors of basidiomycete<br />
species (Pöggeler <strong>and</strong> Kück 2001;<br />
Kim <strong>and</strong> Borkovich 2004). In N. crassa, mutation<br />
of the putative pheromone receptor gene, pre-1,<br />
results in female sterility, although the mutants<br />
are morphologically normal <strong>and</strong> form female<br />
reproductive structures; hyphal fusion in the<br />
pre-1 mutant is normal (Kim <strong>and</strong> Borkovich<br />
2004). Genome sequence analysis of the N. crassa<br />
genome has revealed a large number of additional














