- Page 2 and 3:
The Mycota Edited by K. Esser
- Page 4 and 5:
The Mycota A Comprehensive Treatise
- Page 6 and 7:
Karl Esser (born 1924) is retired P
- Page 8 and 9:
VIII Series Preface Class: Saccharo
- Page 10 and 11:
Addendum to the Series Preface In e
- Page 12 and 13:
XIV Volume Preface to the First Edi
- Page 14 and 15:
XVI Volume Preface to the Second Ed
- Page 16 and 17:
XVIII Contents Reproductive Process
- Page 18 and 19:
XX List of Contributors André Flei
- Page 20 and 21:
Vegetative Processes and Growth
- Page 22 and 23:
4 K.J. Boyce and A. Andrianopoulos
- Page 24 and 25:
6 K.J. Boyce and A. Andrianopoulos
- Page 26 and 27:
8 K.J. Boyce and A. Andrianopoulos
- Page 28 and 29:
10 K.J. Boyce and A. Andrianopoulos
- Page 30 and 31:
12 K.J. Boyce and A. Andrianopoulos
- Page 32 and 33:
14 K.J. Boyce and A. Andrianopoulos
- Page 34 and 35:
16 K.J. Boyce and A. Andrianopoulos
- Page 36 and 37:
18 K.J. Boyce and A. Andrianopoulos
- Page 38 and 39:
20 K.J. Boyce and A. Andrianopoulos
- Page 40 and 41:
22 L.J. García-Rodríguez et al. I
- Page 42 and 43:
24 L.J. García-Rodríguez et al. F
- Page 44 and 45:
26 L.J. García-Rodríguez et al. 2
- Page 46 and 47:
28 L.J. García-Rodríguez et al. M
- Page 48 and 49:
30 L.J. García-Rodríguez et al. a
- Page 50 and 51:
32 L.J. García-Rodríguez et al. t
- Page 52 and 53:
34 L.J. García-Rodríguez et al. H
- Page 54 and 55:
36 L.J. García-Rodríguez et al. T
- Page 56 and 57:
38 S.D. Harris dle organization tha
- Page 58 and 59:
40 S.D. Harris 2001; Borkovich et a
- Page 60 and 61:
42 S.D. Harris terized in A. nidula
- Page 62 and 63:
44 S.D. Harris astral microtubules
- Page 64 and 65:
46 S.D. Harris entry, and the CDK N
- Page 66 and 67:
48 S.D. Harris and Hamer 1997), the
- Page 68 and 69:
50 S.D. Harris Murray AW (2004) Rec
- Page 70 and 71:
4 Apical Wall Biogenesis J.H. Siets
- Page 72 and 73:
fungi can be regarded as “tube-dw
- Page 74 and 75:
In agreement with an essential role
- Page 76 and 77:
Kopecka and Gabriel 1992). They als
- Page 78 and 79:
nearly all the label was present in
- Page 80 and 81:
ingredients such as wall components
- Page 82 and 83:
in membrane enlargement and exocyto
- Page 84 and 85:
sis occurs, coinciding with a gradi
- Page 86 and 87:
pling of two (1-3)-alpha-glucan seg
- Page 88 and 89:
Sietsma JH, Wessels JGH (1977) Chem
- Page 90 and 91:
5 The Fungal Cell Wall J.P. Latgé
- Page 92 and 93:
polymers (chitosan) and glucuronic
- Page 94 and 95:
Whereas the structural branched β1
- Page 96 and 97:
their structural role in the cell w
- Page 98 and 99:
eight chitin synthases of A. fumiga
- Page 100 and 101:
Characterization of the chs4 mutant
- Page 102 and 103:
Fig. 5.8. Experimental data and hyp
- Page 104 and 105:
Fig. 5.9. Elongation of the mannan
- Page 106 and 107:
end of linear β1,3 glucans, and tr
- Page 108 and 109:
Fig. 5.12. Signal transduction in f
- Page 110 and 111:
shock,lowosmolarityaswellasotherfac
- Page 112 and 113:
ditions. Accordingly, enzymes and r
- Page 114 and 115:
Calonge TM, Arellano M, Coll PM, Pe
- Page 116 and 117:
Hiura N, Nakajima T, Matsuda K (198
- Page 118 and 119:
Martin-Yken H, Dagkessamanskaia A,
- Page 120 and 121:
Saporito-Irwin SM, Birse CE, Sypher
- Page 122 and 123:
6 Septation and Cytokinesis in Fung
- Page 124 and 125:
Septation in Fungi 107 Table. 6.1.
- Page 126 and 127:
Fig. 6.1. Selection of a cell divis
- Page 128 and 129:
Calderone, Chap. 5, this volume, an
- Page 130 and 131:
permissive temperature, multiple se
- Page 132 and 133:
eports suggested such a function fo
- Page 134 and 135:
understood mechanism of mitotic exi
- Page 136 and 137:
Implication in cytokinesis in Sacch
- Page 138 and 139:
Trinci APJ, Morris NR (1979) Morpho
- Page 140 and 141:
124 N.L. Glass and A. Fleissner ter
- Page 142 and 143:
126 N.L. Glass and A. Fleissner 188
- Page 144 and 145:
128 N.L. Glass and A. Fleissner Fig
- Page 146 and 147:
130 N.L. Glass and A. Fleissner sub
- Page 148 and 149:
132 N.L. Glass and A. Fleissner dur
- Page 150 and 151:
134 N.L. Glass and A. Fleissner G1
- Page 152 and 153:
136 N.L. Glass and A. Fleissner Bri
- Page 154 and 155:
138 N.L. Glass and A. Fleissner Mat
- Page 156 and 157:
8 Heterogenic Incompatibility in Fu
- Page 158 and 159:
Fungal Heterogenic Incompatibility
- Page 160 and 161:
B. Genetic Control The genetic back
- Page 162 and 163:
active phenotype het-s. The neutral
- Page 164 and 165:
Table. 8.1. (continued) Fungal Hete
- Page 166 and 167:
is a complex genetic trait controll
- Page 168 and 169:
indicate how speciation may be init
- Page 170 and 171:
ility controlled by multiple allele
- Page 172 and 173:
dependonthematingtypegenes,wasobser
- Page 174 and 175:
6. Relation with Histo-Incompatibil
- Page 176 and 177:
Semi-Incompatibilität. Z Indukt Ab
- Page 178 and 179:
Micali CO, Smith ML (2003) On the i
- Page 180 and 181:
Vilgalys RJ, Miller OK (1987) Matin
- Page 182 and 183:
168 B.C.K. Lu (Esser et al. 1980; K
- Page 184 and 185:
170 B.C.K. Lu enterthePCDpathway,wi
- Page 186 and 187:
172 B.C.K. Lu et al. 2004). It is l
- Page 188 and 189:
174 B.C.K. Lu (MMP), through ruptur
- Page 190 and 191:
176 B.C.K. Lu Fig. 9.1. Effects of
- Page 192 and 193:
178 B.C.K. Lu drial fission during
- Page 194 and 195:
180 B.C.K. Lu Although the cytologi
- Page 196 and 197:
182 B.C.K. Lu be arrested at diffus
- Page 198 and 199:
184 B.C.K. Lu Harris MH, Thompson C
- Page 200 and 201:
186 B.C.K. Lu oxygen species, in Kl
- Page 202 and 203:
10 Senescence and Longevity H.D. Os
- Page 204 and 205:
the amplification of plDNA is not a
- Page 206 and 207:
GRISEA is an orthologue of the yeas
- Page 208 and 209:
Fig. 10.3. Copper delivery to the c
- Page 210 and 211:
have been identified, for example,
- Page 212 and 213:
Kück U, Stahl U, Esser K (1981) Pl
- Page 214 and 215:
Signals in Growth and Development
- Page 216 and 217:
204 U. Ugalde II. Germination The p
- Page 218 and 219:
206 U. Ugalde Fig. 11.2.A-C Drawing
- Page 220 and 221:
208 U. Ugalde duction (Schimmel et
- Page 222 and 223:
210 U. Ugalde position, possibly in
- Page 224 and 225:
212 U. Ugalde Champe SP, Rao P, Cha
- Page 226 and 227:
12 Pheromone Action in the Fungal G
- Page 228 and 229:
sibly due to displacement of the hy
- Page 230 and 231:
all these compounds, the B-derivate
- Page 232 and 233:
shows the same activity in M. muced
- Page 234 and 235:
C. Oomycota In the non-mycotan phyl
- Page 236 and 237:
following sequence of events has be
- Page 238 and 239:
the standard Mendelian segregation
- Page 240 and 241:
Elliott CG, Knights BA (1981) Uptak
- Page 242 and 243:
constitutively transcribed but its
- Page 244 and 245:
234 L.M. Corrochano and P. Galland
- Page 246 and 247:
236 L.M. Corrochano and P. Galland
- Page 248 and 249:
238 L.M. Corrochano and P. Galland
- Page 250 and 251:
240 L.M. Corrochano and P. Galland
- Page 252 and 253:
242 L.M. Corrochano and P. Galland
- Page 254 and 255:
244 L.M. Corrochano and P. Galland
- Page 256 and 257:
246 L.M. Corrochano and P. Galland
- Page 258 and 259:
248 L.M. Corrochano and P. Galland
- Page 260 and 261:
250 L.M. Corrochano and P. Galland
- Page 262 and 263:
252 L.M. Corrochano and P. Galland
- Page 264 and 265:
254 L.M. Corrochano and P. Galland
- Page 266 and 267:
256 L.M. Corrochano and P. Galland
- Page 268 and 269:
258 L.M. Corrochano and P. Galland
- Page 270 and 271:
Reproductive Processes
- Page 272 and 273:
264 R. Fischer and U. Kües 2. Chla
- Page 274 and 275:
266 R. Fischer and U. Kües resourc
- Page 276 and 277:
268 R. Fischer and U. Kües ular le
- Page 278 and 279:
270 R. Fischer and U. Kües pathway
- Page 280 and 281:
272 R. Fischer and U. Kües and con
- Page 282 and 283:
274 R. Fischer and U. Kües Timberl
- Page 284 and 285:
276 R. Fischer and U. Kües PsiB le
- Page 286 and 287:
278 R. Fischer and U. Kües Table.
- Page 288 and 289:
280 R. Fischer and U. Kües tein ki
- Page 290 and 291:
282 R. Fischer and U. Kües nitroge
- Page 292 and 293:
284 R. Fischer and U. Kües tion, t
- Page 294 and 295:
286 R. Fischer and U. Kües Busch S
- Page 296 and 297:
288 R. Fischer and U. Kües Jeffs L
- Page 298 and 299:
290 R. Fischer and U. Kües Pöggel
- Page 300 and 301:
292 R. Fischer and U. Kües Yamashi
- Page 302 and 303:
294 R. Debuchy and B.G. Turgeon tha
- Page 304 and 305:
296 R. Debuchy and B.G. Turgeon loc
- Page 306 and 307:
298 R. Debuchy and B.G. Turgeon Tab
- Page 308 and 309:
300 R. Debuchy and B.G. Turgeon Fig
- Page 310 and 311:
302 R. Debuchy and B.G. Turgeon mai
- Page 312 and 313:
304 R. Debuchy and B.G. Turgeon mol
- Page 314 and 315:
306 R. Debuchy and B.G. Turgeon gen
- Page 316 and 317:
308 R. Debuchy and B.G. Turgeon int
- Page 318 and 319:
310 R. Debuchy and B.G. Turgeon Fig
- Page 320 and 321:
312 R. Debuchy and B.G. Turgeon pro
- Page 322 and 323:
314 R. Debuchy and B.G. Turgeon B.
- Page 324 and 325:
316 R. Debuchy and B.G. Turgeon 2.
- Page 326 and 327:
318 R. Debuchy and B.G. Turgeon in
- Page 328 and 329:
320 R. Debuchy and B.G. Turgeon be
- Page 330 and 331:
322 R. Debuchy and B.G. Turgeon Lee
- Page 332 and 333:
16 Fruiting-Body Development in Asc
- Page 334 and 335:
phae originating from the base of a
- Page 336 and 337:
Table. 16.1. (continued) Fruiting B
- Page 338 and 339:
(Raju 1992). When male-sterile muta
- Page 340 and 341:
1. nsd (never in sexual development
- Page 342 and 343:
egular intervals; both effects requ
- Page 344 and 345:
the balance between sexual and asex
- Page 346 and 347:
ductionofeitherpheromoneisdirectlyc
- Page 348 and 349:
(Catlett et al. 2003). Eleven genes
- Page 350 and 351:
negative mutation of KREV-1 resulte
- Page 352 and 353:
through MAP kinase modules to cell-
- Page 354 and 355:
The fact that fruiting-body formati
- Page 356 and 357:
Balestrini R, Mainieri D, Soragni E
- Page 358 and 359:
point mutation of the beta subunit
- Page 360 and 361:
Mitchell TK, Dean RA (1995) The cAM
- Page 362 and 363:
YamashiroCT,EbboleDJ,LeeBU,BrownRE,
- Page 364 and 365:
358 L.A. Casselton and M.P. Challen
- Page 366 and 367:
360 L.A. Casselton and M.P. Challen
- Page 368 and 369:
362 L.A. Casselton and M.P. Challen
- Page 370 and 371:
364 L.A. Casselton and M.P. Challen
- Page 372 and 373:
366 L.A. Casselton and M.P. Challen
- Page 374 and 375:
368 L.A. Casselton and M.P. Challen
- Page 376 and 377:
370 L.A. Casselton and M.P. Challen
- Page 378 and 379: 372 L.A. Casselton and M.P. Challen
- Page 380 and 381: 374 L.A. Casselton and M.P. Challen
- Page 382 and 383: 376 M. Feldbrügge et al. different
- Page 384 and 385: 378 M. Feldbrügge et al. Fig. 18.3
- Page 386 and 387: 380 M. Feldbrügge et al. et al. 20
- Page 388 and 389: 382 M. Feldbrügge et al. for unbia
- Page 390 and 391: 384 M. Feldbrügge et al. In U. may
- Page 392 and 393: 386 M. Feldbrügge et al. Neurospor
- Page 394 and 395: 388 M. Feldbrügge et al. Bernards
- Page 396 and 397: 390 M. Feldbrügge et al. O’Donne
- Page 398 and 399: 19 The Emergence of Fruiting Bodies
- Page 400 and 401: 2003; Kües et al. 2004) are the fi
- Page 402 and 403: (Manachère 1988), C. cinereus (Tsu
- Page 404 and 405: emergence of fruiting bodies. In th
- Page 406 and 407: Rather, development is arrested in
- Page 408 and 409: 10 nm thick is highly insoluble and
- Page 410 and 411: it has been suggested that hydropho
- Page 412 and 413: expression in the stipe suggests th
- Page 414 and 415: Cooper DNW, Boulianne RP, Charlton
- Page 416 and 417: Lu BC (1974) Meiosis in Coprinus. R
- Page 418 and 419: Takagi Y, Katayose Y, Shishido K (1
- Page 420 and 421: 20 Meiosis in Mycelial Fungi D. Zic
- Page 422 and 423: Fig. 20.1. A-E Diagrammatic represe
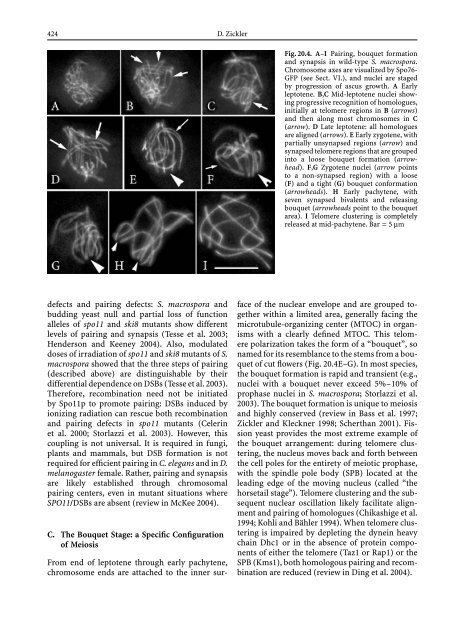
- Page 424 and 425: etween dispersed DNA repeats. In N.
- Page 426 and 427: Fig. 20.2. Meiotic recombination. S
- Page 430 and 431: An important component of the bouqu
- Page 432 and 433: observed when the mammal LE compone
- Page 434 and 435: A. Chromosome and Sister-Chromatid
- Page 436 and 437: ascus plus ascospore morphogenesis
- Page 438 and 439: syndrome)discoveredasageneinvolvedi
- Page 440 and 441: Kitajima TS, Kawashima SA, Watanabe
- Page 442 and 443: Rossignol J-L, Faugeron G (1995) MI
- Page 444 and 445: Biosystematic Index Absidia glauca
- Page 446 and 447: violaceum 153 Microsporum 149 Mimos
- Page 448 and 449: Subject Index ABC transporter 382 a
- Page 450 and 451: fluffy 270, 276 formin 8, 10, 13, 1
- Page 452 and 453: MAT protein interaction 308, 315 ma
- Page 454: sporangiophore 234, 242, 246-252, 2