Kalakasvatus ja kalade tervishoid.indd - Eesti Maaülikool
Kalakasvatus ja kalade tervishoid.indd - Eesti Maaülikool
Kalakasvatus ja kalade tervishoid.indd - Eesti Maaülikool

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Kalade geneetika <strong>ja</strong> aretus<br />
Arvutame:<br />
• selektsioonidiferents S = (604 + 692)/2 – 454 = 194 g,<br />
• oodatav valikuedu R = 194 × 0,3 = 58 g,<br />
• järgmise põlvkonna oodatav 1+ keskmine mass = 454 + 58 = 512 g.<br />
Mida suurem on tunnuse h2 , seda efektiivsem on selektsioon <strong>ja</strong> suurem oodatav valikuedu<br />
(kui h2 = 1, siis R = S; kui aga h2 = 0, siis R = 0!). Tuleb aga silmas pidada, et h2 väärtused ei ole põlvkonniti püsivad, vaid sõltuvad alleelisageduste <strong>ja</strong> keskkonna (<strong>kalade</strong>l<br />
eelkõige temperatuur) muutustest. Seega tuleb hinnata h2 keskkonnas, kus tehakse valikut,<br />
ning see kehtib ainult konkreetse populatsiooni/tõu kohta. Samas sõltub h2 väärtus<br />
ka kala vanusest tunnuse fenotüübi määramise a<strong>ja</strong>l, mistõttu ei saa näiteks samasuviste<br />
<strong>kalade</strong> kehamassi h2 automaatselt üle kanda kolmesuviste <strong>kalade</strong> kehamassile. Tabelis 27<br />
on toodud näiteid valikuedust kasvukiiruse osas mõnedel kalaliikidel.<br />
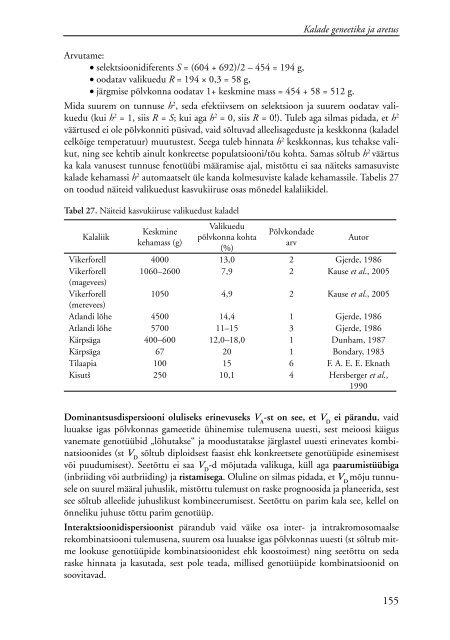
Tabel 27. Näiteid kasvukiiruse valikuedust <strong>kalade</strong>l<br />
Kalaliik<br />
Keskmine<br />
kehamass (g)<br />
Valikuedu<br />
põlvkonna kohta<br />
(%)<br />
Põlvkondade<br />
arv<br />
Autor<br />
Vikerforell 4000 13,0 2 Gjerde, 1986<br />
Vikerforell<br />
(magevees)<br />
1060–2600 7,9 2 Kause et al., 2005<br />
Vikerforell<br />
(merevees)<br />
1050 4,9 2 Kause et al., 2005<br />
Atlandi lõhe 4500 14,4 1 Gjerde, 1986<br />
Atlandi lõhe 5700 11–15 3 Gjerde, 1986<br />
Kärpsäga 400–600 12,0–18,0 1 Dunham, 1987<br />
Kärpsäga 67 20 1 Bondary, 1983<br />
Tilaapia 100 15 6 F. A. E. E. Eknath<br />
Kisutš 250 10,1 4 Hersberger et al.,<br />
1990<br />
Dominantsusdispersiooni oluliseks erinevuseks V -st on see, et V ei pärandu, vaid<br />
A D<br />
luuakse igas põlvkonnas gameetide ühinemise tulemusena uuesti, sest meioosi käigus<br />
vanemate genotüübid „lõhutakse“ <strong>ja</strong> moodustatakse järglastel uuesti erinevates kombinatsioonides<br />
(st V sõltub diploidsest faasist ehk konkreetsete genotüüpide esinemisest<br />
D<br />
või puudumisest). Seetõttu ei saa V -d mõjutada valikuga, küll aga paarumistüübiga<br />
D<br />
(inbriiding või autbriiding) <strong>ja</strong> ristamisega. Oluline on silmas pidada, et V mõju tunnu-<br />
D<br />
sele on suurel määral juhuslik, mistõttu tulemust on raske prognoosida <strong>ja</strong> planee rida, sest<br />
see sõltub alleelide juhuslikust kombineerumisest. Seetõttu on parim kala see, kellel on<br />
õnneliku juhuse tõttu parim genotüüp.<br />
Interaktsioonidispersioonist pärandub vaid väike osa inter- <strong>ja</strong> intrakromosomaalse<br />
rekombinatsiooni tulemusena, suurem osa luuakse igas põlvkonnas uuesti (st sõltub mitme<br />
lookuse genotüüpide kombinatsioonidest ehk koostoimest) ning seetõttu on seda<br />
raske hinnata <strong>ja</strong> kasutada, sest pole teada, millised genotüüpide kombinatsioonid on<br />
soovitavad.<br />
155