

Literature review: Impact of Chilean needle grass ... - Weeds Australia
Literature review: Impact of Chilean needle grass ... - Weeds Australia
Literature review: Impact of Chilean needle grass ... - Weeds Australia

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
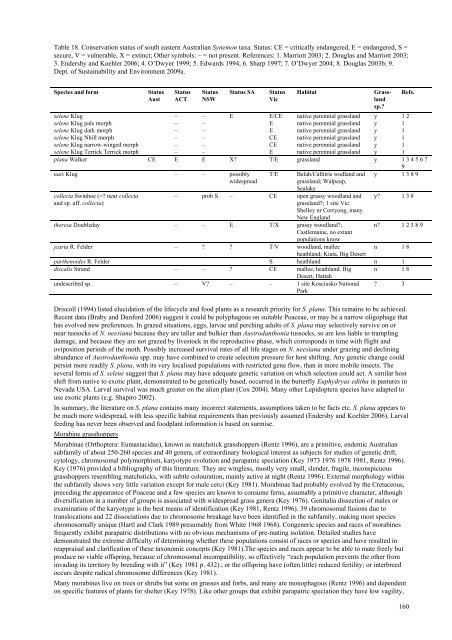
Table 18. Conservation status <strong>of</strong> south eastern <strong>Australia</strong>n Synemon taxa. Status: CE = critically endangered, E = endangered, S =<br />
secure, V = vulnerable, X = extinct; Other symbols: – = not present. References: 1. Marriott 2003; 2. Douglas and Marriott 2003;<br />
3. Endersby and Koehler 2006; 4. O’Dwyer 1999; 5. Edwards 1994, 6. Sharp 1997; 7. O’Dwyer 2004; 8. Douglas 2003b; 9.<br />
Dept. <strong>of</strong> Sustainability and Environment 2009a.<br />
Species and form<br />
Status<br />
Aust<br />
Status<br />
ACT<br />
Status<br />
NSW<br />
Status SA<br />
Status<br />
Vic<br />
Habitat<br />
selene Klug – – E E/CE native perennial <strong>grass</strong>land y 1 2<br />
selene Klug pale morph – – E native perennial <strong>grass</strong>land y 1<br />
selene Klug dark morph – – E native perennial <strong>grass</strong>land y 1<br />
selene Klug Nhill morph – – CE native perennial <strong>grass</strong>land y 1<br />
selene Klug narrow-winged morph – – CE native perennial <strong>grass</strong>land y 1<br />
selene Klug Terrick Terrick morph – – E native perennial <strong>grass</strong>land y 1<br />
plana Walker CE E E X? T/E <strong>grass</strong>land y 1 3 4 5 6 7<br />
9<br />
nais Klug – – possibly<br />
widespread<br />
collecta Swinhoe (=? near collecta<br />
and sp. aff. collecta)<br />
T/E<br />
Belah/Callitris wodland and<br />
<strong>grass</strong>land; Walpeup,<br />
Sealake<br />
– prob S – CE open <strong>grass</strong>y woodland and<br />
<strong>grass</strong>land?; 1 site Vic:<br />
Shelley nr Corryong, many<br />
New England<br />
Grassland<br />
sp.?<br />
Refs.<br />
y 1 3 8 9<br />
y? 1 3 8<br />
theresa Doubleday – – E T/X <strong>grass</strong>y woodland?;<br />
n? 1 2 3 8 9<br />
Castlemaine, no extant<br />
populations know<br />
jcaria R. Felder – ? ? T/V woodland, mallee<br />
n 1 8<br />
heathland; Kiata, Big Desert<br />
parthenoides R. Felder S heathland n 1<br />
discalis Strand – – ? CE mallee, heathland. Big n 1 8<br />
Desert, Hattah<br />
undescribed sp. – V? – – 1 site Kosciusko National<br />
Park<br />
? 3<br />
Driscoll (1994) listed elucidation <strong>of</strong> the lifecycle and food plants as a research priority for S. plana. This remains to be achieved.<br />
Recent data (Braby and Dunford 2006) suggest it could be polyphagous on suitable Poaceae, or may be a narrow oligophage that<br />
has evolved new preferences. In grazed situations, eggs, larvae and perching adults <strong>of</strong> S. plana may selectively survive on or<br />
near tussocks <strong>of</strong> N. neesiana because they are taller and bulkier than Austrodanthonia tussocks, so are less liable to trampling<br />
damage, and because they are not grazed by livestock in the reproductive phase, which corresponds in time with flight and<br />
oviposition periods <strong>of</strong> the moth. Possibly increased survival rates <strong>of</strong> all life stages on N. neesiana under grazing and declining<br />
abundance <strong>of</strong> Austrodanthonia spp. may have combined to create selection pressure for host shifting. Any genetic change could<br />
persist more readily S. plana, with its very localised populations with restricted gene flow, than in more mobile insects. The<br />
several forms <strong>of</strong> S. selene suggest that S. plana may have adequate genetic variation on which selection could act. A similar host<br />
shift from native to exotic plant, demonstrated to be genetically based, occurred in the butterfly Euphydryas editha in pastures in<br />
Nevada USA. Larval survival was much greater on the alien plant (Cox 2004). Many other Lepidoptera species have adapted to<br />
use exotic plants (e.g. Shapiro 2002).<br />
In summary, the literature on S. plana contains many incorrect statements, assumptions taken to be facts etc. S. plana appears to<br />
be much more widespread, with less specific habitat requirements than previously assumed (Endersby and Koehler 2006). Larval<br />
feeding has never been observed and foodplant information is based on surmise.<br />
Morabine <strong>grass</strong>hoppers<br />
Morabinae (Orthoptera: Eumastacidae), known as matchstick <strong>grass</strong>hoppers (Rentz 1996), are a primitive, endemic <strong>Australia</strong>n<br />
subfamily <strong>of</strong> about 250-260 species and 40 genera, <strong>of</strong> extraordinary biological interest as subjects for studies <strong>of</strong> genetic drift,<br />
cytology, chromosomal polymorphism, karyotype evolution and parapatric speciation (Key 1973 1976 1978 1981, Rentz 1996).<br />
Key (1976) provided a bibliography <strong>of</strong> this literature. They are wingless, mostly very small, slender, fragile, inconspicuous<br />
<strong>grass</strong>hoppers resembling matchsticks, with subtle colouration, mainly active at night (Rentz 1996). External morphology within<br />
the subfamily shows very little variation except for male cerci (Key 1981). Morabinae had probably evolved by the Cretaceous,<br />
preceding the appearance <strong>of</strong> Poaceae and a few species are known to consume ferns, assumably a primitive character, although<br />
diversification in a number <strong>of</strong> groups is associated with widespread <strong>grass</strong> genera (Key 1976). Genitalia dissection <strong>of</strong> males or<br />
examination <strong>of</strong> the karyotype is the best means <strong>of</strong> identification (Key 1981, Rentz 1996). 39 chromosomal fusions due to<br />
translocations and 22 dissociations due to chromosome breakage have been identified in the subfamily, making most species<br />
chromosomally unique (Hartl and Clark 1989 presumably from White 1968 1968). Congeneric species and races <strong>of</strong> morabines<br />
frequently exhibit parapatric distributions with no obvious mechanisms <strong>of</strong> pre-mating isolation. Detailed studies have<br />
demonstrated the extreme difficulty <strong>of</strong> determining whether these populations consist <strong>of</strong> races or species and have resulted in<br />
reappraisal and clarification <strong>of</strong> these taxonomic concepts (Key 1981).The species and races appear to be able to mate freely but<br />
produce no viable <strong>of</strong>fspring, because <strong>of</strong> chromosomal incompatibility, so effectively “each population prevents the other from<br />
invading its territory by breeding with it” (Key 1981 p. 432).; or the <strong>of</strong>fspring have (<strong>of</strong>ten little) reduced fertility; or interbreed<br />
occurs despite radical chromosome differences (Key 1981).<br />
Many morabines live on trees or shrubs but some on <strong>grass</strong>es and forbs, and many are monophagous (Rentz 1996) and dependent<br />
on specific features <strong>of</strong> plants for shelter (Key 1978). Like other groups that exhibit parapatric speciation they have low vagility,<br />
160














