

Proceedings of the Third International Conference on Invasive ...
Proceedings of the Third International Conference on Invasive ...
Proceedings of the Third International Conference on Invasive ...
- No tags were found...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
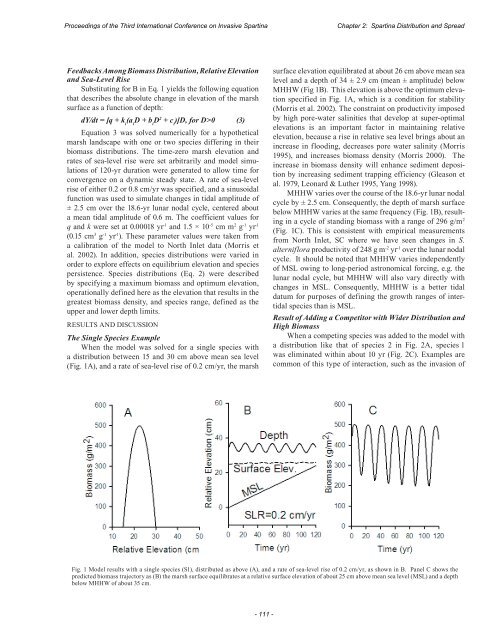
<str<strong>on</strong>g>Proceedings</str<strong>on</strong>g> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>Third</str<strong>on</strong>g> <str<strong>on</strong>g>Internati<strong>on</strong>al</str<strong>on</strong>g> <str<strong>on</strong>g>C<strong>on</strong>ference</str<strong>on</strong>g> <strong>on</strong> <strong>Invasive</strong> SpartinaChapter 2: Spartina Distributi<strong>on</strong> and SpreadFeedbacks Am<strong>on</strong>g Biomass Distributi<strong>on</strong>, Relative Elevati<strong>on</strong>and Sea-Level RiseSubstituting for B in Eq. 1 yields <str<strong>on</strong>g>the</str<strong>on</strong>g> following equati<strong>on</strong>that describes <str<strong>on</strong>g>the</str<strong>on</strong>g> absolute change in elevati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> marshsurface as a functi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> depth:dY/dt = [q + k i(a iD + b iD 2 + c i)]D, for D>0 (3)Equati<strong>on</strong> 3 was solved numerically for a hypo<str<strong>on</strong>g>the</str<strong>on</strong>g>ticalmarsh landscape with <strong>on</strong>e or two species differing in <str<strong>on</strong>g>the</str<strong>on</strong>g>irbiomass distributi<strong>on</strong>s. The time-zero marsh elevati<strong>on</strong> andrates <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise were set arbitrarily and model simulati<strong>on</strong>s<str<strong>on</strong>g>of</str<strong>on</strong>g> 120-yr durati<strong>on</strong> were generated to allow time forc<strong>on</strong>vergence <strong>on</strong> a dynamic steady state. A rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-levelrise <str<strong>on</strong>g>of</str<strong>on</strong>g> ei<str<strong>on</strong>g>the</str<strong>on</strong>g>r 0.2 or 0.8 cm/yr was specified, and a sinusoidalfuncti<strong>on</strong> was used to simulate changes in tidal amplitude <str<strong>on</strong>g>of</str<strong>on</strong>g>± 2.5 cm over <str<strong>on</strong>g>the</str<strong>on</strong>g> 18.6-yr lunar nodal cycle, centered abouta mean tidal amplitude <str<strong>on</strong>g>of</str<strong>on</strong>g> 0.6 m. The coefficient values forq and k were set at 0.00018 yr -1 and 1.5 × 10 -5 cm m 2 g -1 yr -1(0.15 cm 3 g -1 yr -1 ). These parameter values were taken froma calibrati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> model to North Inlet data (Morris etal. 2002). In additi<strong>on</strong>, species distributi<strong>on</strong>s were varied inorder to explore effects <strong>on</strong> equilibrium elevati<strong>on</strong> and speciespersistence. Species distributi<strong>on</strong>s (Eq. 2) were describedby specifying a maximum biomass and optimum elevati<strong>on</strong>,operati<strong>on</strong>ally defined here as <str<strong>on</strong>g>the</str<strong>on</strong>g> elevati<strong>on</strong> that results in <str<strong>on</strong>g>the</str<strong>on</strong>g>greatest biomass density, and species range, defined as <str<strong>on</strong>g>the</str<strong>on</strong>g>upper and lower depth limits.RESULTS AND DISCUSSIONThe Single Species ExampleWhen <str<strong>on</strong>g>the</str<strong>on</strong>g> model was solved for a single species witha distributi<strong>on</strong> between 15 and 30 cm above mean sea level(Fig. 1A), and a rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise <str<strong>on</strong>g>of</str<strong>on</strong>g> 0.2 cm/yr, <str<strong>on</strong>g>the</str<strong>on</strong>g> marshsurface elevati<strong>on</strong> equilibrated at about 26 cm above mean sealevel and a depth <str<strong>on</strong>g>of</str<strong>on</strong>g> 34 ± 2.9 cm (mean ± amplitude) belowMHHW (Fig 1B). This elevati<strong>on</strong> is above <str<strong>on</strong>g>the</str<strong>on</strong>g> optimum elevati<strong>on</strong>specified in Fig. 1A, which is a c<strong>on</strong>diti<strong>on</strong> for stability(Morris et al. 2002). The c<strong>on</strong>straint <strong>on</strong> productivity imposedby high pore-water salinities that develop at super-optimalelevati<strong>on</strong>s is an important factor in maintaining relativeelevati<strong>on</strong>, because a rise in relative sea level brings about anincrease in flooding, decreases pore water salinity (Morris1995), and increases biomass density (Morris 2000). Theincrease in biomass density will enhance sediment depositi<strong>on</strong>by increasing sediment trapping efficiency (Gleas<strong>on</strong> etal. 1979, Le<strong>on</strong>ard & Lu<str<strong>on</strong>g>the</str<strong>on</strong>g>r 1995, Yang 1998).MHHW varies over <str<strong>on</strong>g>the</str<strong>on</strong>g> course <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> 18.6-yr lunar nodalcycle by ± 2.5 cm. C<strong>on</strong>sequently, <str<strong>on</strong>g>the</str<strong>on</strong>g> depth <str<strong>on</strong>g>of</str<strong>on</strong>g> marsh surfacebelow MHHW varies at <str<strong>on</strong>g>the</str<strong>on</strong>g> same frequency (Fig. 1B), resultingin a cycle <str<strong>on</strong>g>of</str<strong>on</strong>g> standing biomass with a range <str<strong>on</strong>g>of</str<strong>on</strong>g> 296 g/m 2(Fig. 1C). This is c<strong>on</strong>sistent with empirical measurementsfrom North Inlet, SC where we have seen changes in S.alterniflora productivity <str<strong>on</strong>g>of</str<strong>on</strong>g> 248 g m -2 yr -1 over <str<strong>on</strong>g>the</str<strong>on</strong>g> lunar nodalcycle. It should be noted that MHHW varies independently<str<strong>on</strong>g>of</str<strong>on</strong>g> MSL owing to l<strong>on</strong>g-period astr<strong>on</strong>omical forcing, e.g. <str<strong>on</strong>g>the</str<strong>on</strong>g>lunar nodal cycle, but MHHW will also vary directly withchanges in MSL. C<strong>on</strong>sequently, MHHW is a better tidaldatum for purposes <str<strong>on</strong>g>of</str<strong>on</strong>g> defining <str<strong>on</strong>g>the</str<strong>on</strong>g> growth ranges <str<strong>on</strong>g>of</str<strong>on</strong>g> intertidalspecies than is MSL.Result <str<strong>on</strong>g>of</str<strong>on</strong>g> Adding a Competitor with Wider Distributi<strong>on</strong> andHigh BiomassWhen a competing species was added to <str<strong>on</strong>g>the</str<strong>on</strong>g> model witha distributi<strong>on</strong> like that <str<strong>on</strong>g>of</str<strong>on</strong>g> species 2 in Fig. 2A, species 1was eliminated within about 10 yr (Fig. 2C). Examples arecomm<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> this type <str<strong>on</strong>g>of</str<strong>on</strong>g> interacti<strong>on</strong>, such as <str<strong>on</strong>g>the</str<strong>on</strong>g> invasi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g>Fig. 1 Model results with a single species (S1), distributed as above (A), and a rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise <str<strong>on</strong>g>of</str<strong>on</strong>g> 0.2 cm/yr, as shown in B. Panel C shows <str<strong>on</strong>g>the</str<strong>on</strong>g>predicted biomass trajectory as (B) <str<strong>on</strong>g>the</str<strong>on</strong>g> marsh surface equilibrates at a relative surface elevati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> about 25 cm above mean sea level (MSL) and a depthbelow MHHW <str<strong>on</strong>g>of</str<strong>on</strong>g> about 35 cm.- 111 -




