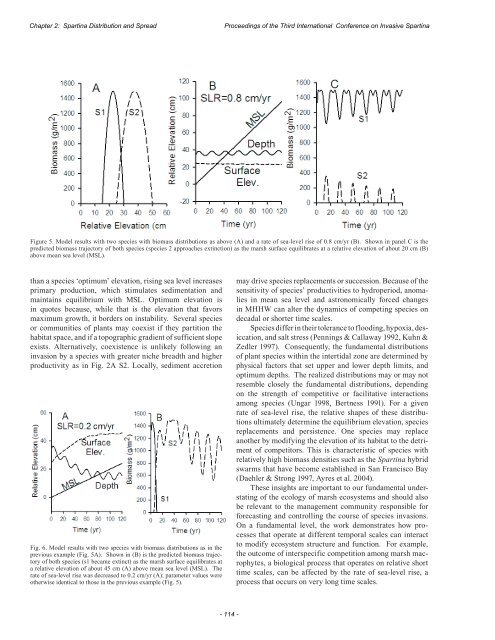
<str<strong>on</strong>g>Proceedings</str<strong>on</strong>g> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>Third</str<strong>on</strong>g> <str<strong>on</strong>g>Internati<strong>on</strong>al</str<strong>on</strong>g> <str<strong>on</strong>g>C<strong>on</strong>ference</str<strong>on</strong>g> <strong>on</strong> <strong>Invasive</strong> SpartinaChapter 2: Spartina Distributi<strong>on</strong> and SpreadFig.4. Model results with <strong>on</strong>e species with a biomass distributi<strong>on</strong> as above (A) and a rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise <str<strong>on</strong>g>of</str<strong>on</strong>g> 0.8 cm/yr. Shown in panel C is <str<strong>on</strong>g>the</str<strong>on</strong>g> predictedbiomass trajectory <str<strong>on</strong>g>of</str<strong>on</strong>g> species 2 as (B) <str<strong>on</strong>g>the</str<strong>on</strong>g> marsh surface equilibrates at an elevati<strong>on</strong> below MSL and outside <str<strong>on</strong>g>of</str<strong>on</strong>g> species 2’s range. Parameter values wereo<str<strong>on</strong>g>the</str<strong>on</strong>g>rwise identical to those in <str<strong>on</strong>g>the</str<strong>on</strong>g> previous examples (Fig. 2 and 3).The biomass <str<strong>on</strong>g>of</str<strong>on</strong>g> species 2 varied with depth at <str<strong>on</strong>g>the</str<strong>on</strong>g> frequency<str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> lunar nodal cycle, but 180º out-<str<strong>on</strong>g>of</str<strong>on</strong>g>-phase, whereas<str<strong>on</strong>g>the</str<strong>on</strong>g> lunar nodal cycle and biomass <str<strong>on</strong>g>of</str<strong>on</strong>g> species 1 were in phase(Fig. 3B). The biomass <str<strong>on</strong>g>of</str<strong>on</strong>g> species 2 decreased with a rise inMHHW, i.e. as equilibrium depth increased, because <str<strong>on</strong>g>the</str<strong>on</strong>g>marsh surface elevati<strong>on</strong>, 28 cm, was suboptimal for growth.C<strong>on</strong>versely, <str<strong>on</strong>g>the</str<strong>on</strong>g> biomass <str<strong>on</strong>g>of</str<strong>on</strong>g> species 1 increased with a rise inMHHW because surface elevati<strong>on</strong> was super-optimal for itsgrowth. Thus, species can coexist <str<strong>on</strong>g>the</str<strong>on</strong>g>oretically when <str<strong>on</strong>g>the</str<strong>on</strong>g>reis a periodic change in flood durati<strong>on</strong> and when <strong>on</strong>e speciesresp<strong>on</strong>ds to <str<strong>on</strong>g>the</str<strong>on</strong>g> change positively and <str<strong>on</strong>g>the</str<strong>on</strong>g> o<str<strong>on</strong>g>the</str<strong>on</strong>g>r negatively.This is <strong>on</strong>e <str<strong>on</strong>g>of</str<strong>on</strong>g> several c<strong>on</strong>diti<strong>on</strong>s that promote a facultativeinteracti<strong>on</strong> between species.Facultative interacti<strong>on</strong>s am<strong>on</strong>g marsh macrophytesinvolving ameliorati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> soil salinity have been described(Bertness & Ewanchuk 2002), and simulati<strong>on</strong> results shownhere suggest that biogeomorphological interacti<strong>on</strong>s couldalso be facultative. When two species with overlapping distributi<strong>on</strong>s(Fig. 2A) were present, and <str<strong>on</strong>g>the</str<strong>on</strong>g> rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-levelrise was raised to 0.8 cm/yr, <str<strong>on</strong>g>the</str<strong>on</strong>g> resulting interacti<strong>on</strong> couldbe described as facultative (Fig. 3), because nei<str<strong>on</strong>g>the</str<strong>on</strong>g>r speciespersists in <str<strong>on</strong>g>the</str<strong>on</strong>g> absence <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> o<str<strong>on</strong>g>the</str<strong>on</strong>g>r (Fig. 4). When species 1was removed from <str<strong>on</strong>g>the</str<strong>on</strong>g> simulati<strong>on</strong>, <str<strong>on</strong>g>the</str<strong>on</strong>g> equilibrium elevati<strong>on</strong>quickly dropped below MSL and below <str<strong>on</strong>g>the</str<strong>on</strong>g> lower limit forspecies 2. Species 1 suffered <str<strong>on</strong>g>the</str<strong>on</strong>g> same fate when species 2was removed. However, at a low rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise, 0.2cm/yr, species 1 did not survive (Fig. 2). Thus, <str<strong>on</strong>g>the</str<strong>on</strong>g> outcome<str<strong>on</strong>g>of</str<strong>on</strong>g> competiti<strong>on</strong> and <str<strong>on</strong>g>the</str<strong>on</strong>g> emergence <str<strong>on</strong>g>of</str<strong>on</strong>g> facultative behaviordepend <strong>on</strong> <str<strong>on</strong>g>the</str<strong>on</strong>g> rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise.The Rate <str<strong>on</strong>g>of</str<strong>on</strong>g> Sea-Level Rise and Alternative Stable StatesIt can also be dem<strong>on</strong>strated that <str<strong>on</strong>g>the</str<strong>on</strong>g> marsh will movetoward alternative stable states or habitat preempti<strong>on</strong> by <strong>on</strong>especies or ano<str<strong>on</strong>g>the</str<strong>on</strong>g>r, depending <strong>on</strong> <str<strong>on</strong>g>the</str<strong>on</strong>g> rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level riseand <str<strong>on</strong>g>the</str<strong>on</strong>g> species’ biomass distributi<strong>on</strong>s. When <str<strong>on</strong>g>the</str<strong>on</strong>g> rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sealevelrise was 0.8 cm/yr and <str<strong>on</strong>g>the</str<strong>on</strong>g> distributi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> species 1 wasmodified by raising its maximum biomass to equal that <str<strong>on</strong>g>of</str<strong>on</strong>g>species 2 (Fig. 5A), <strong>on</strong>ly species 1 persisted throughout <str<strong>on</strong>g>the</str<strong>on</strong>g>simulati<strong>on</strong>, while species 2 was intermittent (Fig. 5C). Themarsh surface equilibrated at an elevati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> about 23 cm(Fig. 5B), which was at <str<strong>on</strong>g>the</str<strong>on</strong>g> lower limit <str<strong>on</strong>g>of</str<strong>on</strong>g> species 2 and above<str<strong>on</strong>g>the</str<strong>on</strong>g> optimum elevati<strong>on</strong> for species 1 (Fig. 5A). Keeping <str<strong>on</strong>g>the</str<strong>on</strong>g>species distributi<strong>on</strong>s as in Fig. 5A and lowering <str<strong>on</strong>g>the</str<strong>on</strong>g> rate <str<strong>on</strong>g>of</str<strong>on</strong>g>sea-level rise to 0.2 cm/yr resulted in a different outcome. Atthis lower rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise, <strong>on</strong>ly species 2 persisted (Fig.6C). The marsh surface equilibrated at an elevati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> about46 cm (Fig. 6A), which is above <str<strong>on</strong>g>the</str<strong>on</strong>g> upper limit <str<strong>on</strong>g>of</str<strong>on</strong>g> species 1and above <str<strong>on</strong>g>the</str<strong>on</strong>g> optimum <str<strong>on</strong>g>of</str<strong>on</strong>g> species 2 (Fig. 5A).CONCLUSIONSThis paper describes a <str<strong>on</strong>g>the</str<strong>on</strong>g>ory <str<strong>on</strong>g>of</str<strong>on</strong>g> biogeomorphology thataddresses how intertidal macrophytes can modify landscapeelevati<strong>on</strong> and affect <str<strong>on</strong>g>the</str<strong>on</strong>g> outcomes <str<strong>on</strong>g>of</str<strong>on</strong>g> species interacti<strong>on</strong>sby means <str<strong>on</strong>g>of</str<strong>on</strong>g> vertical geomorphological displacement.Competiti<strong>on</strong> by geomorphogical displacement is indirectand is a functi<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> fundamental and realized niches thatdescribe species biomass distributi<strong>on</strong>s in <str<strong>on</strong>g>the</str<strong>on</strong>g> tidal frame. Theoutcomes <str<strong>on</strong>g>of</str<strong>on</strong>g> competitive interacti<strong>on</strong>s are a great deal morecomplex than described here when <str<strong>on</strong>g>the</str<strong>on</strong>g>re are direct interferencesor facultative interacti<strong>on</strong>s <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> sort described byBertness and Ewanchuk (2002). These types <str<strong>on</strong>g>of</str<strong>on</strong>g> interacti<strong>on</strong>scan modify a species’ realized distributi<strong>on</strong>s dynamically andwould result in truly complex behavior.Marsh primary producti<strong>on</strong> and standing biomass aresensitive to hydroperiod and have positive effects <strong>on</strong> sedimentaccreti<strong>on</strong>. In that part <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> tidal frame that is higher- 113 -
Chapter 2: Spartina Distributi<strong>on</strong> and Spread<str<strong>on</strong>g>Proceedings</str<strong>on</strong>g> <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> <str<strong>on</strong>g>Third</str<strong>on</strong>g> <str<strong>on</strong>g>Internati<strong>on</strong>al</str<strong>on</strong>g> <str<strong>on</strong>g>C<strong>on</strong>ference</str<strong>on</strong>g> <strong>on</strong> <strong>Invasive</strong> SpartinaFigure 5. Model results with two species with biomass distributi<strong>on</strong>s as above (A) and a rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise <str<strong>on</strong>g>of</str<strong>on</strong>g> 0.8 cm/yr (B). Shown in panel C is <str<strong>on</strong>g>the</str<strong>on</strong>g>predicted biomass trajectory <str<strong>on</strong>g>of</str<strong>on</strong>g> both species (species 2 approaches extincti<strong>on</strong>) as <str<strong>on</strong>g>the</str<strong>on</strong>g> marsh surface equilibrates at a relative elevati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> about 20 cm (B)above mean sea level (MSL).than a species ‘optimum’ elevati<strong>on</strong>, rising sea level increasesprimary producti<strong>on</strong>, which stimulates sedimentati<strong>on</strong> andmaintains equilibrium with MSL. Optimum elevati<strong>on</strong> isin quotes because, while that is <str<strong>on</strong>g>the</str<strong>on</strong>g> elevati<strong>on</strong> that favorsmaximum growth, it borders <strong>on</strong> instability. Several speciesor communities <str<strong>on</strong>g>of</str<strong>on</strong>g> plants may coexist if <str<strong>on</strong>g>the</str<strong>on</strong>g>y partiti<strong>on</strong> <str<strong>on</strong>g>the</str<strong>on</strong>g>habitat space, and if a topographic gradient <str<strong>on</strong>g>of</str<strong>on</strong>g> sufficient slopeexists. Alternatively, coexistence is unlikely following aninvasi<strong>on</strong> by a species with greater niche breadth and higherproductivity as in Fig. 2A S2. Locally, sediment accreti<strong>on</strong>Fig. 6. Model results with two species with biomass distributi<strong>on</strong>s as in <str<strong>on</strong>g>the</str<strong>on</strong>g>previous example (Fig. 5A). Shown in (B) is <str<strong>on</strong>g>the</str<strong>on</strong>g> predicted biomass trajectory<str<strong>on</strong>g>of</str<strong>on</strong>g> both species (s1 became extinct) as <str<strong>on</strong>g>the</str<strong>on</strong>g> marsh surface equilibrates ata relative elevati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> about 45 cm (A) above mean sea level (MSL). Therate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise was decreased to 0.2 cm/yr (A); parameter values wereo<str<strong>on</strong>g>the</str<strong>on</strong>g>rwise identical to those in <str<strong>on</strong>g>the</str<strong>on</strong>g> previous example (Fig. 5).may drive species replacements or successi<strong>on</strong>. Because <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g>sensitivity <str<strong>on</strong>g>of</str<strong>on</strong>g> species’ productivities to hydroperiod, anomaliesin mean sea level and astr<strong>on</strong>omically forced changesin MHHW can alter <str<strong>on</strong>g>the</str<strong>on</strong>g> dynamics <str<strong>on</strong>g>of</str<strong>on</strong>g> competing species <strong>on</strong>decadal or shorter time scales.Species differ in <str<strong>on</strong>g>the</str<strong>on</strong>g>ir tolerance to flooding, hypoxia, desiccati<strong>on</strong>,and salt stress (Pennings & Callaway 1992, Kuhn &Zedler 1997). C<strong>on</strong>sequently, <str<strong>on</strong>g>the</str<strong>on</strong>g> fundamental distributi<strong>on</strong>s<str<strong>on</strong>g>of</str<strong>on</strong>g> plant species within <str<strong>on</strong>g>the</str<strong>on</strong>g> intertidal z<strong>on</strong>e are determined byphysical factors that set upper and lower depth limits, andoptimum depths. The realized distributi<strong>on</strong>s may or may notresemble closely <str<strong>on</strong>g>the</str<strong>on</strong>g> fundamental distributi<strong>on</strong>s, depending<strong>on</strong> <str<strong>on</strong>g>the</str<strong>on</strong>g> strength <str<strong>on</strong>g>of</str<strong>on</strong>g> competitive or facilitative interacti<strong>on</strong>sam<strong>on</strong>g species (Ungar 1998, Bertness 1991). For a givenrate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise, <str<strong>on</strong>g>the</str<strong>on</strong>g> relative shapes <str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g>se distributi<strong>on</strong>sultimately determine <str<strong>on</strong>g>the</str<strong>on</strong>g> equilibrium elevati<strong>on</strong>, speciesreplacements and persistence. One species may replaceano<str<strong>on</strong>g>the</str<strong>on</strong>g>r by modifying <str<strong>on</strong>g>the</str<strong>on</strong>g> elevati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> its habitat to <str<strong>on</strong>g>the</str<strong>on</strong>g> detriment<str<strong>on</strong>g>of</str<strong>on</strong>g> competitors. This is characteristic <str<strong>on</strong>g>of</str<strong>on</strong>g> species withrelatively high biomass densities such as <str<strong>on</strong>g>the</str<strong>on</strong>g> Spartina hybridswarms that have become established in San Francisco Bay(Daehler & Str<strong>on</strong>g 1997, Ayres et al. 2004).These insights are important to our fundamental understating<str<strong>on</strong>g>of</str<strong>on</strong>g> <str<strong>on</strong>g>the</str<strong>on</strong>g> ecology <str<strong>on</strong>g>of</str<strong>on</strong>g> marsh ecosystems and should alsobe relevant to <str<strong>on</strong>g>the</str<strong>on</strong>g> management community resp<strong>on</strong>sible forforecasting and c<strong>on</strong>trolling <str<strong>on</strong>g>the</str<strong>on</strong>g> course <str<strong>on</strong>g>of</str<strong>on</strong>g> species invasi<strong>on</strong>s.On a fundamental level, <str<strong>on</strong>g>the</str<strong>on</strong>g> work dem<strong>on</strong>strates how processesthat operate at different temporal scales can interactto modify ecosystem structure and functi<strong>on</strong>. For example,<str<strong>on</strong>g>the</str<strong>on</strong>g> outcome <str<strong>on</strong>g>of</str<strong>on</strong>g> interspecific competiti<strong>on</strong> am<strong>on</strong>g marsh macrophytes,a biological process that operates <strong>on</strong> relative shorttime scales, can be affected by <str<strong>on</strong>g>the</str<strong>on</strong>g> rate <str<strong>on</strong>g>of</str<strong>on</strong>g> sea-level rise, aprocess that occurs <strong>on</strong> very l<strong>on</strong>g time scales.- 114 -