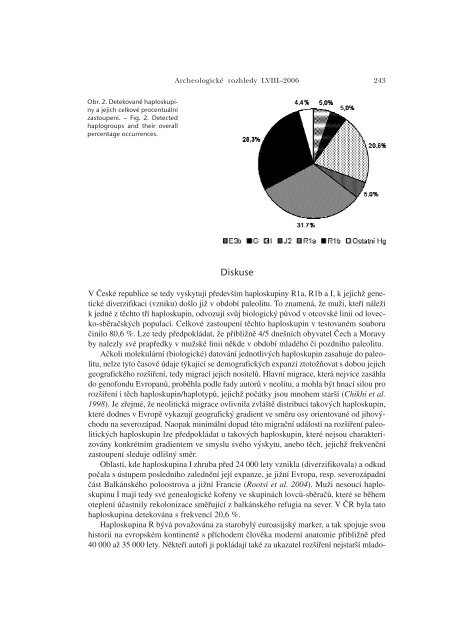
242KRÁâMAROVÁ –BRUCHOVÁ –âERN¯ –BRDIâKA: Podíl „paleolitick˘ch“ …LokalitaHaploskupina – počet (%)E3b G I J2 R1a R1b Ostatní HgKlatovy – 3 (10,0) 8 (26,7) 2 (6,7) 11 (36,7) 5 (16,7) 1 (3,3)Písek – 2 (6,7) 8 (26,7) 1 (3,3) 8 (26,7) 9 (30,0) 2 (6,7)Jindřichův Hradec 4 (13,3) 3 (10,0) 4 (13,3) 1 (3,3) 9 (30,0) 9 (30,0) –Třebíč 3 (10,0) 1 (3,3) 5 (16,7) 3 (10,0) 8 (26,7) 9 (30,0) 1 (3,3)Vyškov 1 (3,3) – 6 (20,0) 2 (6,7) 7 (23,3) 12 (40,0) 2 (6,7)Vsetín 1 (3,3) – 6 (20,0) – 14 (46,7) 7 (23,3) 2 (6,7)Celkem 9 (5,0) 9 (5,0) 37 (20,6) 9 (5,0) 57 (31,7) 51 (28,3) 8 (4,4)Tab. 1. Detekované haploskupiny a jejich počet (procentuální zastoupení) u všech populačních skupin.– Tab. 1. Detected haplogroups and their numbers (percentage occurrences) in all population groups.genetického názvosloví, který vychází z pravidel Y chromozómového konsorcia z roku 2002 (viz internetovéodkazy).Částečně odlišné složení testovaných polymorfismů umožnilo autorům Semino et al. (2000) charakterizovati jiné haploskupiny v porovnání s naší prací. Např. vzorky vykazující odvozenou alelu v polymorfismuM207 (charakterizuje celou haploskupinu R) byly v původní studii testovány v polymorfismuM17, přičemž byla-li přítomna v tomto markeru odvozená alela, byl příslušný vzorek zařazen do haploskupinyR1a1. Naopak pokud byla zaznamenána v polymorfismu M17 alela původní, byly dané Y chromozomyklasifikovány jako R*(R1a1). Na druhou stranu námi detekované vzorky mající odvozenou aleluv polymorfismu M207 byly dále sledovány ve dvou zcela odlišných polymorfismech SRY10831.b a P25.První z nich určuje haploskupinu R1a, jež v sobě mimo jiné zahrnuje i hierarchicky níže postavenou haploskupinuR1a1, druhý marker určuje její sesterskou haploskupinu R1b. Jediný vzorek našeho souboru,který vykazoval odvozenou alelu v polymorfismu M207, nikoli však v následujících polymorfismechSRY10831.b a P25, byl zařazen do haploskupiny R*(xR1a, R1b). Za účelem srovnání výsledků obou pracíjsme vytvořili společnou kategorii označenou R*(xR1a), do které byly začleněny všechny Y chromozomyklasifikované jako R1b a R*(xR1a, R1b). Její frekvence (28,9 %) pak byla srovnána s frekvencíodpovídající haploskupiny R*(xR1a1) (35,6 %) publikovanou ve studii Semino et al. 2000. Analogickyvýskyt námi detekované haploskupiny R1a (31,7 %) byl konfrontován se zastoupením haploskupinyR1a1 (26,7 %).VýsledkyV testovaném souboru 180 vzorků české populace bylo detekováno deset haploskupin,z nichž šest (E3b, G, I, J2, R1a a R1b) dosahovalo frekvence rovnu 5 % či vyšší. K ostatnímčtyřem minoritním haploskupinám (F* či K*, H, P*, R* (xR1a, R1b)), které se vyskytovalys relativní četností nižší než 5 %, náleželo pouze 8 vzorků. Výsledky shrnutév tab. 1 ukazují, že nejméně haploskupin (n=5) bylo pozorováno u populačního vzorkuze Vsetína, naopak nejvíce (n=7) jich bylo zaznamenáno mezi vzorky z Třebíče a Vyškova.Celkové četnosti jednotlivých haploskupin detekovaných v této práci dokumentujeobr. 2. Nejvyššího výskytu dosahují haploskupiny R1a (31,7 %), R1b (28,3 %) a I (20,6 %).Mezi relativně početné patřily také haploskupiny E3b, G a J2, shodně se v <strong>ČR</strong> vyskytujícís frekvencí 5 %.
<strong>Archeologické</strong> <strong>rozhledy</strong> LVIII–<strong>2006</strong> 243Obr. 2. Detekované haploskupinya jejich celkové procentuálnízastoupení. – Fig. 2. Detectedhaplogroups and their overallpercentage occurrences.DiskuseV České republice se tedy vyskytují především haploskupiny R1a, R1b a I, k jejichž genetickédiverzifikaci (vzniku) došlo již v období paleolitu. To znamená, že muži, kteří náležík jedné z těchto tří haploskupin, odvozují svůj biologický původ v otcovské linii od lovecko-sběračskýchpopulací. Celkové zastoupení těchto haploskupin v testovaném souboručinilo 80,6 %. Lze tedy předpokládat, že přibližně 4/5 dnešních obyvatel Čech a Moravyby nalezly své prapředky v mužské linii někde v období mladého či pozdního paleolitu.Ačkoli molekulární (biologické) datování jednotlivých haploskupin zasahuje do paleolitu,nelze tyto časové údaje týkající se demografických expanzí ztotožňovat s dobou jejichgeografického rozšíření, tedy migrací jejich nositelů. Hlavní migrace, která nejvíce zasáhlado genofondu Evropanů, proběhla podle řady autorů v neolitu, a mohla být hnací silou prorozšíření i těch haploskupin/haplotypů, jejichž počátky jsou mnohem starší (Chikhi et al.1998). Je zřejmé, že neolitická migrace ovlivnila zvláště distribuci takových haploskupin,které dodnes v Evropě vykazují geografický gradient ve směru osy orientované od jihovýchoduna severozápad. Naopak minimální dopad této migrační události na rozšíření paleolitickýchhaploskupin lze předpokládat u takových haploskupin, které nejsou charakterizoványkonkrétním gradientem ve smyslu svého výskytu, anebo těch, jejichž frekvenčnízastoupení sleduje odlišný směr.Oblastí, kde haploskupina I zhruba před 24 000 lety vznikla (diverzifikovala) a odkudpočala s ústupem posledního zalednění její expanze, je jižní Evropa, resp. severozápadníčást Balkánského poloostrova a jižní Francie (Rootsi et al. 2004). Muži nesoucí haploskupinuI mají tedy své genealogické kořeny ve skupinách lovců-sběračů, které se běhemoteplení účastnily rekolonizace směřující z balkánského refugia na sever. V <strong>ČR</strong> byla tatohaploskupina detekována s frekvencí 20,6 %.Haploskupina R bývá považována za starobylý euroasijský marker, a tak spojuje svouhistorii na evropském kontinentě s příchodem člověka moderní anatomie přibližně před40 000 až 35 000 lety. Někteří autoři ji pokládají také za ukazatel rozšíření nejstarší mlado-