Informe Técnico Gestión de pH en el vino de calidad
Informe Técnico Gestión de pH en el vino de calidad
Informe Técnico Gestión de pH en el vino de calidad

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
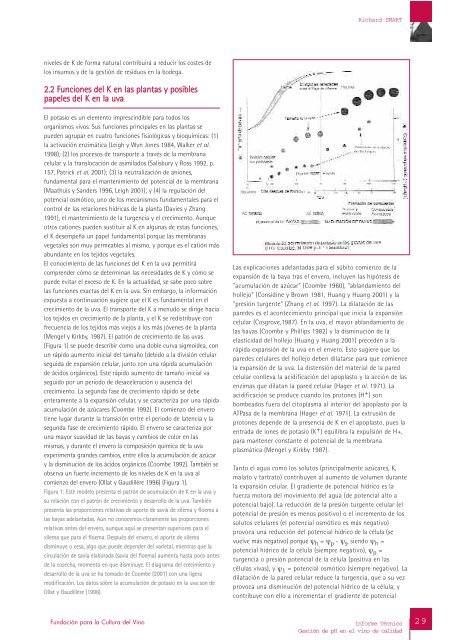
Richard SMARTniv<strong>el</strong>es <strong>de</strong> K <strong>de</strong> forma natural contribuirá a reducir los costes <strong>de</strong>los insumos y <strong>de</strong> la gestión <strong>de</strong> residuos <strong>en</strong> la bo<strong>de</strong>ga.2.2 Funciones <strong>de</strong>l K <strong>en</strong> las plantas y posiblespap<strong>el</strong>es <strong>de</strong>l K <strong>en</strong> la uvaEl potasio es un <strong>el</strong>em<strong>en</strong>to imprescindible para todos losorganismos vivos. Sus funciones principales <strong>en</strong> las plantas sepue<strong>de</strong>n agrupar <strong>en</strong> cuatro funciones fisiológicas y bioquímicas: (1)la activación <strong>en</strong>zimática (Leigh y Wyn Jones 1984, Walker et al.1998); (2) los procesos <strong>de</strong> transporte a través <strong>de</strong> la membranac<strong>el</strong>ular y la translocación <strong>de</strong> asimilados (Salisbury y Ross 1992, p.157, Patrick et al. 2001); (3) la neutralización <strong>de</strong> aniones,fundam<strong>en</strong>tal para <strong>el</strong> mant<strong>en</strong>imi<strong>en</strong>to <strong>de</strong>l pot<strong>en</strong>cial <strong>de</strong> la membrana(Maathuis y San<strong>de</strong>rs 1996, Leigh 2001); y (4) la regulación <strong>de</strong>lpot<strong>en</strong>cial osmótico, uno <strong>de</strong> los mecanismos fundam<strong>en</strong>tales para <strong>el</strong>control <strong>de</strong> las r<strong>el</strong>aciones hídricas <strong>de</strong> la planta (Davies y Zhang1991), <strong>el</strong> mant<strong>en</strong>imi<strong>en</strong>to <strong>de</strong> la turg<strong>en</strong>cia y <strong>el</strong> crecimi<strong>en</strong>to. Aunqueotros cationes pue<strong>de</strong>n sustituir al K <strong>en</strong> algunas <strong>de</strong> estas funciones,<strong>el</strong> K <strong>de</strong>sempeña un pap<strong>el</strong> fundam<strong>en</strong>tal porque las membranasvegetales son muy permeables al mismo, y porque es <strong>el</strong> catión másabundante <strong>en</strong> los tejidos vegetales.El conocimi<strong>en</strong>to <strong>de</strong> las funciones <strong>de</strong>l K <strong>en</strong> la uva permitirácompr<strong>en</strong><strong>de</strong>r cómo se <strong>de</strong>terminan las necesida<strong>de</strong>s <strong>de</strong> K y cómo sepue<strong>de</strong> evitar <strong>el</strong> exceso <strong>de</strong> K. En la actualidad, se sabe poco sobr<strong>el</strong>as funciones exactas <strong>de</strong>l K <strong>en</strong> la uva. Sin embargo, la informaciónexpuesta a continuación sugiere que <strong>el</strong> K es fundam<strong>en</strong>tal <strong>en</strong> <strong>el</strong>crecimi<strong>en</strong>to <strong>de</strong> la uva. El transporte <strong>de</strong>l K a m<strong>en</strong>udo se dirige hacialos tejidos <strong>en</strong> crecimi<strong>en</strong>to <strong>de</strong> la planta, y <strong>el</strong> K se redistribuye confrecu<strong>en</strong>cia <strong>de</strong> los tejidos más viejos a los más jóv<strong>en</strong>es <strong>de</strong> la planta(M<strong>en</strong>g<strong>el</strong> y Kirkby, 1987). El patrón <strong>de</strong> crecimi<strong>en</strong>to <strong>de</strong> las uvas(Figura 1) se pue<strong>de</strong> <strong>de</strong>scribir como una doble curva sigmoi<strong>de</strong>a, conun rápido aum<strong>en</strong>to inicial <strong>de</strong>l tamaño (<strong>de</strong>bido a la división c<strong>el</strong>ularseguida <strong>de</strong> expansión c<strong>el</strong>ular, junto con una rápida acumulación<strong>de</strong> ácidos orgánicos). Este rápido aum<strong>en</strong>to <strong>de</strong> tamaño inicial vaseguido por un periodo <strong>de</strong> <strong>de</strong>sac<strong>el</strong>eración o aus<strong>en</strong>cia <strong>de</strong>lcrecimi<strong>en</strong>to. La segunda fase <strong>de</strong> crecimi<strong>en</strong>to rápido se <strong>de</strong>be<strong>en</strong>teram<strong>en</strong>te a la expansión c<strong>el</strong>ular, y se caracteriza por una rápidaacumulación <strong>de</strong> azúcares (Coombe 1992). El comi<strong>en</strong>zo <strong>de</strong>l <strong>en</strong>veroti<strong>en</strong>e lugar durante la transición <strong>en</strong>tre <strong>el</strong> periodo <strong>de</strong> lat<strong>en</strong>cia y lasegunda fase <strong>de</strong> crecimi<strong>en</strong>to rápido. El <strong>en</strong>vero se caracteriza poruna mayor suavidad <strong>de</strong> las bayas y cambios <strong>de</strong> color <strong>en</strong> lasmismas, y durante <strong>el</strong> <strong>en</strong>vero la composición química <strong>de</strong> la uvaexperim<strong>en</strong>ta gran<strong>de</strong>s cambios, <strong>en</strong>tre <strong>el</strong>los la acumulación <strong>de</strong> azúcary la disminución <strong>de</strong> los ácidos orgánicos (Coombe 1992). También seobserva un fuerte increm<strong>en</strong>to <strong>de</strong> los niv<strong>el</strong>es <strong>de</strong> K <strong>en</strong> la uva alcomi<strong>en</strong>zo <strong>de</strong>l <strong>en</strong>vero (Ollat y Gaudillère 1996) (Figura 1).Figura 1: Este mo<strong>de</strong>lo pres<strong>en</strong>ta <strong>el</strong> patrón <strong>de</strong> acumulación <strong>de</strong> K <strong>en</strong> la uva ysu r<strong>el</strong>ación con <strong>el</strong> patrón <strong>de</strong> crecimi<strong>en</strong>to y <strong>de</strong>sarrollo <strong>de</strong> la uva. Tambiénpres<strong>en</strong>ta las proporciones r<strong>el</strong>ativas <strong>de</strong> aporte <strong>de</strong> savia <strong>de</strong> xilema y floema alas bayas a<strong>de</strong>lantadas. Aún no conocemos claram<strong>en</strong>te las proporcionesr<strong>el</strong>ativas antes <strong>de</strong>l <strong>en</strong>vero, aunque aquí se pres<strong>en</strong>tan superiores para <strong>el</strong>xilema que para <strong>el</strong> floema. Después <strong>de</strong>l <strong>en</strong>vero, <strong>el</strong> aporte <strong>de</strong> xilemadisminuye o cesa, algo que pue<strong>de</strong> <strong>de</strong>p<strong>en</strong><strong>de</strong>r <strong>de</strong>l varietal, mi<strong>en</strong>tras que lacirculación <strong>de</strong> savia <strong>el</strong>aborada (savia <strong>de</strong>l floema) aum<strong>en</strong>ta hasta poco antes<strong>de</strong> la cosecha, mom<strong>en</strong>to <strong>en</strong> que disminuye. El diagrama <strong>de</strong>l crecimi<strong>en</strong>to y<strong>de</strong>sarrollo <strong>de</strong> la uva se ha tomado <strong>de</strong> Coombe (2001) con una ligeramodificación. Los datos sobre la acumulación <strong>de</strong> potasio <strong>en</strong> la uva son <strong>de</strong>Ollat y Gaudillère (1996).Las explicaciones a<strong>de</strong>lantadas para <strong>el</strong> súbito comi<strong>en</strong>zo <strong>de</strong> laexpansión <strong>de</strong> la baya tras <strong>el</strong> <strong>en</strong>vero, incluy<strong>en</strong> las hipótesis <strong>de</strong>"acumulación <strong>de</strong> azúcar" (Coombe 1960), "ablandami<strong>en</strong>to <strong>de</strong>lhollejo" (Considine y Brown 1981, Huang y Huang 2001) y la"presión turg<strong>en</strong>te" (Zhang et al. 1997). La dilatación <strong>de</strong> laspare<strong>de</strong>s es <strong>el</strong> acontecimi<strong>en</strong>to principal que inicia la expansiónc<strong>el</strong>ular (Cosgrove,1987). En la uva, <strong>el</strong> mayor ablandami<strong>en</strong>to <strong>de</strong>las bayas (Coombe y Phillips 1982) y la disminución <strong>de</strong> la<strong>el</strong>asticidad <strong>de</strong>l hollejo (Huang y Huang 2001) prece<strong>de</strong>n a larápida expansión <strong>de</strong> la uva <strong>en</strong> <strong>el</strong> <strong>en</strong>vero. Esto sugiere que laspare<strong>de</strong>s c<strong>el</strong>ulares <strong>de</strong>l hollejo <strong>de</strong>b<strong>en</strong> dilatarse para que comi<strong>en</strong>c<strong>el</strong>a expansión <strong>de</strong> la uva. La dist<strong>en</strong>sión <strong>de</strong>l material <strong>de</strong> la paredc<strong>el</strong>ular conlleva la acidificación <strong>de</strong>l apoplasto y la acción <strong>de</strong> las<strong>en</strong>zimas que dilatan la pared c<strong>el</strong>ular (Hager et al. 1971). Laacidificación se produce cuando los protones (H + ) sonbombeados fuera <strong>de</strong>l citoplasma al interior <strong>de</strong>l apoplasto por laATPasa <strong>de</strong> la membrana (Hager et al. 1971). La extrusión <strong>de</strong>protones <strong>de</strong>p<strong>en</strong><strong>de</strong> <strong>de</strong> la pres<strong>en</strong>cia <strong>de</strong> K <strong>en</strong> <strong>el</strong> apoplasto, pues la<strong>en</strong>trada <strong>de</strong> iones <strong>de</strong> potasio (K + ) equilibra la expulsión <strong>de</strong> H+,para mant<strong>en</strong>er constante <strong>el</strong> pot<strong>en</strong>cial <strong>de</strong> la membranaplasmática (M<strong>en</strong>g<strong>el</strong> y Kirkby 1987).Tanto <strong>el</strong> agua como los solutos (principalm<strong>en</strong>te azúcares, K,malato y tartrato) contribuy<strong>en</strong> al aum<strong>en</strong>to <strong>de</strong> volum<strong>en</strong> durant<strong>el</strong>a expansión c<strong>el</strong>ular. El gradi<strong>en</strong>te <strong>de</strong> pot<strong>en</strong>cial hídrico es lafuerza motora <strong>de</strong>l movimi<strong>en</strong>to <strong>de</strong>l agua (<strong>de</strong> pot<strong>en</strong>cial alto apot<strong>en</strong>cial bajo). La reducción <strong>de</strong> la presión turg<strong>en</strong>te c<strong>el</strong>ular (<strong>el</strong>pot<strong>en</strong>cial <strong>de</strong> presión es m<strong>en</strong>os positivo) o <strong>el</strong> increm<strong>en</strong>to <strong>de</strong> lossolutos c<strong>el</strong>ulares (<strong>el</strong> pot<strong>en</strong>cial osmótico es más negativo)provoca una reducción <strong>de</strong>l pot<strong>en</strong>cial hídrico <strong>de</strong> la célula (sevu<strong>el</strong>ve más negativo) porque ψ h = ψ p - ψ s , si<strong>en</strong>do ψ h =pot<strong>en</strong>cial hídrico <strong>de</strong> la célula (siempre negativo), ψ p =turg<strong>en</strong>cia o presión pot<strong>en</strong>cial <strong>de</strong> la célula (positiva <strong>en</strong> lascélulas vivas), y ψ 1 = pot<strong>en</strong>cial osmótico (siempre negativo). Ladilatación <strong>de</strong> la pared c<strong>el</strong>ular reduce la turg<strong>en</strong>cia, que a su vezprovoca una disminución <strong>de</strong>l pot<strong>en</strong>cial hídrico <strong>de</strong> la célula, ycontribuye con <strong>el</strong>lo a increm<strong>en</strong>tar <strong>el</strong> gradi<strong>en</strong>te <strong>de</strong> pot<strong>en</strong>cial29Fundación para la Cultura <strong>de</strong>l Vino<strong>Informe</strong> TécnicoGestión <strong>de</strong> <strong>pH</strong> <strong>en</strong> <strong>el</strong> <strong>vino</strong> <strong>de</strong> <strong>calidad</strong>29